2001 Nature Publishing Group http://neurosci.nature.com brief communications L-type Ca2+ channel-
In sharp contrast to the lack of effect on single IPSCs, blockade
of L-type Ca2+ channels eliminated PTP and turned it into a depres-
mediated short-term
sion lasting for about 80 s (Fig. 2b–d). In nifedipine, the post- tetanus eIPSC1 was reduced to 85.3 ± 11% of control, versus plasticity of GABAergic
129 ± 12% of control before nifedipine was perfused (p < 0.05, n= 4, paired t-test). In diltiazem, the post-tetanus eIPSC1 was 76.3
synapses
± 7.1% of control (Fig. 3b), versus 122 ± 6.4% of control before diltiazem (p < 0.05, n = 4). These findings are consistent with a crit- ical role of L-type Ca2+ channels in the induction of post-tetanic
potentiation of GABAergic IPSCs. In the presence of nifedipine,
the post-tetanic IPSCs showed paired-pulse facilitation, so that the
Department of Neurology, RNRC 3-131, University of California, Los Angeles
eIPSC2:eIPSC1 ratio increased from 0.69 ± 0.06 to 1.07 ± 0.15 (Fig. School of Medicine, 710 Westwood Plaza, Los Angeles, California 90095, USA 3c, p < 0.05, paired t-test). In diltiazem, the tetanus increased the Correspondence should be addressed to I.M. (mody@ucla.edu)
eIPSC2:eIPSC1 ratio from 0.86 ± 0.07 to 1.27 ± 0.25 (p < 0.05). The
osci.nature
L-type channel antagonists had no effect on the progressive tetan-
Published online: 10 September 2001, DOI: 10.1038/nn722
ic depression of the eIPSCs occurring during the 80-pulse stimu-lation at 40 Hz (p > 0.05). Nifedipine and diltiazem also failed to
In the cerebral cortex, the major inhibitory neurotransmitter GABA
block the post-tetanus increase in spontaneous IPSCs. (See the sup-
(γ-aminobutyric acid) is released by GABAergic neurons1 onto
plementary information page of Nature Neuroscience online.)
GABAA and GABAB receptors, and regulates neuronal excitability,
Here we confirm that L-type Ca2+ channels are not involved in
oup http://neur
postsynaptic action potential firing, and dendritic and synaptic inte-
low-frequency synaptic transmission at GABAergic synapses8. How-
gration2. Various interneurons use either N- or P/Q-type Ca2+ chan-
ever, the contribution of L-type channels to synaptic transmission
nels for the Ca2+ influx into their terminals3, whereas L-type Ca2+
became evident when GABAergic synapses were driven by high-
channels are not normally associated with GABA release. In dual
frequency firing in the gamma-frequency range4, which led to PTP. lishing Gr
recordings from hippocampal basket cells and granule cells, we now
The rapid onset of PTP suggests that the presynaptic L-type Ca2+
report that short-term plasticity of GABA release is controlled by
channels are located near the GABAergic terminals. This is sur-
L-type Ca2+ channels at presynaptic firing rates in the gamma-fre-
prising because L-type channels have yet to be anatomically detect-
quency (40 Hz) range4; at these GABAergic synapses, L-type Ca2+
ed at these sights. Given that L-type Ca2+ channel antagonists block
channel antagonists converted post-tetanic potentiation into depres-
PTP but not the ‘late release’ (above and ref. 8), whereas EGTA-AM
sion, identifying L-type Ca2+ channels as important modulators of
blocks ‘late release’ but not PTP7,9, we propose that the L-type Ca2+
channels are coupled to second messenger cascades regulating the
2001 Nature Pub
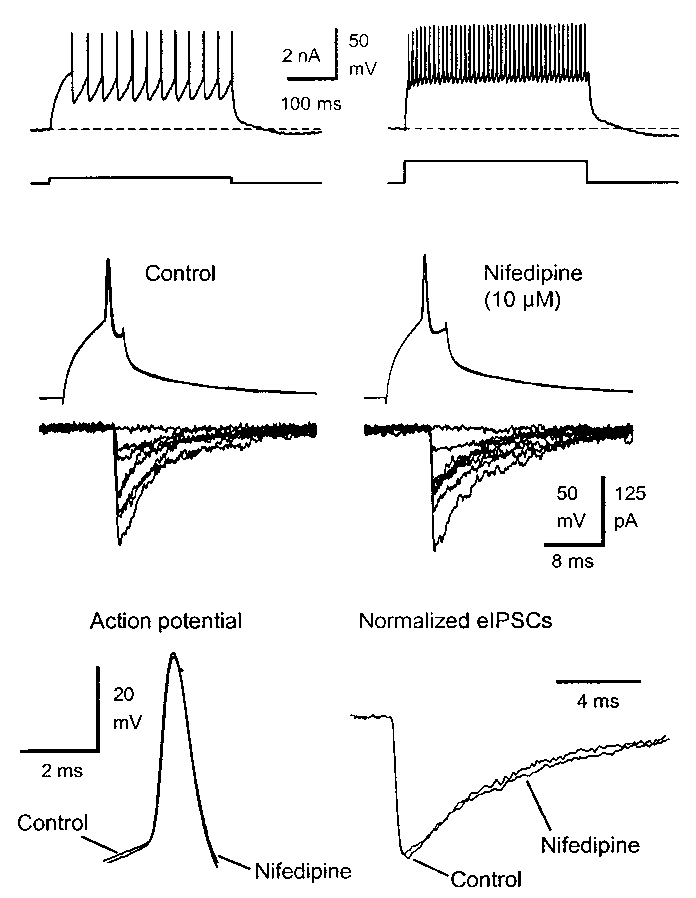
Fast-spiking basket cells (Fig. 1a) were identified on the border
availability of transmitter vesicle pools. L-type Ca2+ channel activ-
between the granule cell layer and the hilus in the rat dentate gyrus
ity could activate calmodulin-mediated processes10,11, which in
in brain slices. In dual recordings from basket cells and granule cellsin 3 mM kynurenic acid, stimulation of the basket cell to fire sin-gle action potentials (APs) evoked a GABAA receptor-mediated
IPSC (eIPSC) in the granule cell (Fig. 1b). Baseline responses were obtained by presynaptic paired-pulse stimulation (50 ms interval) evoking two eIPSCs (eIPSC1 and eIPSC2) every 5 s. The amplitude of eIPSC1 was 139 ± 32 pA (33°C, n = 9) and showed quantal-like variations, including failures (11.1 ± 5.4%). Perfusion of the L-type
Ca2+ channel antagonist nifedipine (10 µM) did not affect thepresynaptic AP (half-width, 0.53 ± 0.06 ms in control versus 0.60 ± 0.08 ms with nifedipine, p > 0.05, n = 4). Nifedipine alsohad no effect on the eIPSC1 amplitude (96 ± 15% of control, p > 0.05, n = 4) or its variance, latency, 10–90% rise time or decaykinetics. Another L-type Ca2+ channel blocker, diltiazem (25 µM),also failed to affect eIPSC1 (amplitude, 105 ± 16% of control, p > 0.05, n = 4) or the presynaptic AP. Miniature IPSCs recordedin granule cells in the presence of CdCl
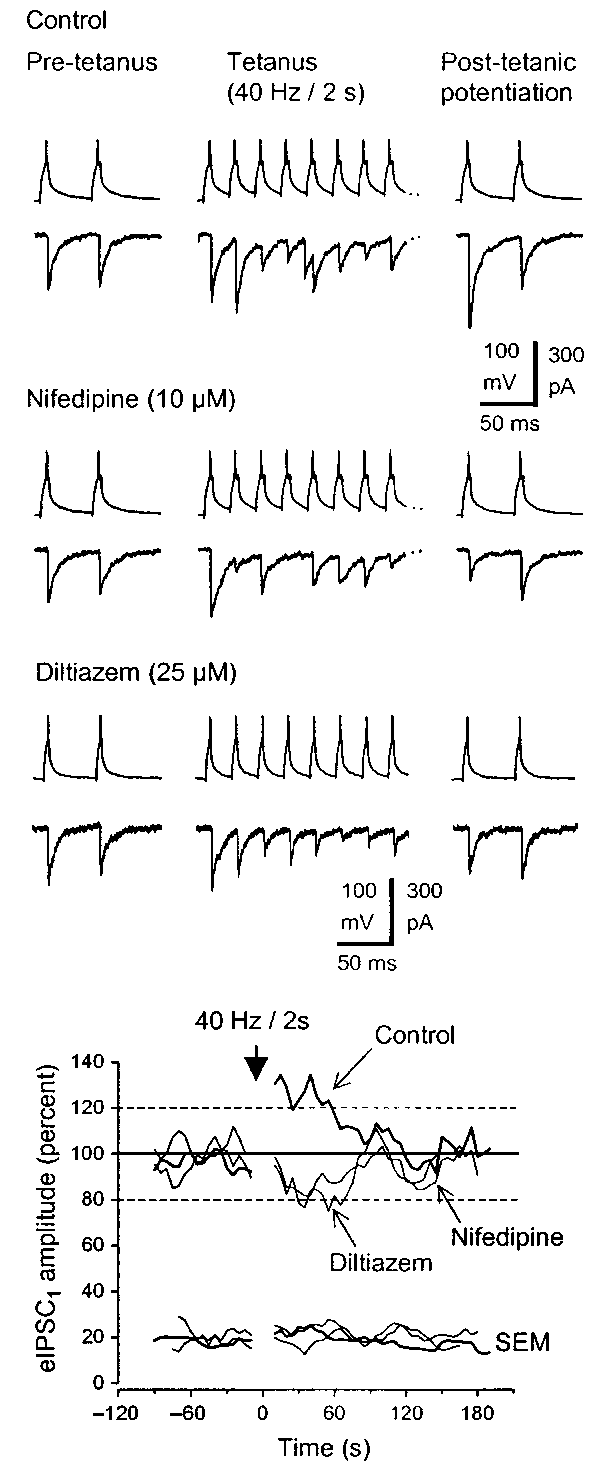
We then examined the short-term plasticity of IPSCs evoked by
high-frequency APs in the range of the normal firing behavior ofbasket cells5. We induced post-tetanic potentiation (PTP) of eIP-SCs by stimulating the basket cells in the gamma-frequency range(40 Hz, 2 s). PTP is a reversible enhancement of transmitter release
Fig 1. Nifedipine did not affect GABAergic IPSCs evoked by low-fre-
that lasts for minutes, and has been observed at many synapses6,7. In
quency basket cell firing. (a) Basket cell firing in the dentate gyrus in rat
brain slices. (b) Dual whole-cell recording from a basket cell and a gran-
5–45 s post-tetanus, p < 0.05, paired t-test) and lasted about 120 s
ule cell (bottom) in 3 mM kynurenic acid. Stimulation of the basket cell
(Fig. 2a and d, n = 12). During this enhancement, paired-pulse
to fire single action potentials evoked short-latency eIPSCs in the gran-
depression of the eIPSCs also increased, consistent with PTP being
ule cell. Consecutive eIPSCs were evoked every 5 s and superimposed. The L-type Ca2+ channel antagonist nifedipine (10 µM) did not affect the
a presynaptic phenomenon. Hence, the eIPSC2:eIPSC1 ratio
eIPSCs. (c) The presynaptic action potential and the eIPSCs shown on
decreased from 0.95 ± 0.06 to 0.84 ± 0.07 (p < 0.05, paired t-test).
an expanded time scale. Their time course was unaffected by nifedipine. nature neuroscience • volume 4 no 10 • october 2001 2001 Nature Publishing Group http://neurosci.nature.com brief communications osci.nature Fig. 3. The post-tetanic eIPSC amplitude and paired-pulse modulation. (a) PTP in a single cell pair. Data were analyzed as indicated by brackets. Pre-tetanic, average of 12 eIPSCs before the tetanus; post-tetanic, aver- age of 9 eIPSC after the tetanus; recovery, average of 12 eIPSCs when the responses had recovered. (b) Post-tetanic eIPSC1 in control (white oup http://neur
bars, n = 12), in nifedipine (n = 4) and in diltiazem (n = 4). The normal- ized eIPSC1 amplitudes showed PTP in control, and depression in nifedipine and diltiazem (p < 0.05). (c) The eIPSC2:eIPSC1 ratios illus-
trating the paired-pulse behavior of the IPSCs. In control, post-tetanic
lishing Gr
IPSCs showed a small increase in paired-pulse depression (white bars, p< 0.05). In the presence of the L-type Ca2+ channel blockers, post-tetanic eIPSCs displayed paired-pulse facilitation (p < 0.05), indicatingthat the release probability had been lowered.
channel-mediated potentiation of GABA release, which may be crit-
2001 Nature Pub
ical for frequency shifts15 in the cerebral cortex. Therefore, L-type
Ca2+ channels could be important in controlling the populationactivity in highly interconnected hippocampal neurons. Fig. 2. Post-tetanic potentiation of IPSCs was blocked by L-type Ca2+ channel antagonists. (a) Paired-pulse stimulation of the basket cell to Note: Supplementary methods are available on the Nature Neuroscience web site
fire APs evoked two IPSCs (eIPSC1 and eIPSC2) every 5 s in the granulecell. After tetanic stimulation of the basket cell (40 Hz, 2 s), eIPSC
(http://neuroscience.nature.com/web_specials.)
played post-tetanic potentiation by 75% and the paired-pulse depres- sion of eIPSC2 increased. (b) Following perfusion of nifedipine, the ACKNOWLEDGEMENTS
pre-tetanic eIPSC1 was unaffected, whereas the post-tetanic eIPSC1 was
This work was supported by NIH grant NS 30549 and the Coelho Endowment to
depressed to 63% of control and eIPSC2 showed paired-pulse facilita-
I.M. K.J. is a Glaxo/Wellcome scholar.
tion. (c) The L-type channel antagonist diltiazem (25 µM) also caused post-tetanic depression. (d) Post-tetanic potentiation of eIPSC1 in con- RECEIVED 2 JULY; ACCEPTED 18 AUGUST 2001
trol (n = 12) compared with post-tetanic depression in nifedipine (n = 4)or diltiazem (n = 4). Tetanic stimulation of the basket cell was deliveredat 0 s. For each trial, eIPSC
1. Freund, T. F. & Buzsaki, G. Hippocampus 6, 347–470 (1996).
2. Miles, R., Toth, K., Gulyas, A. I., Hajos, N. & Freund, T. F. Neuron 16, 815–823
tetanic baseline (100%), plotted and averaged across cells. The plot was
smoothed by a running average of six responses. The standard error for
3. Poncer, J. C., McKinney, R. A., Gahwiler, B. H. & Thompson, S. M. Neuron 18,
each group (also smoothed) is shown below.
4. Tamas, G., Buhl, E. H., Lorincz, A. & Somogyi, P. Nat. Neurosci. 3, 366–371
5. Penttonen, M., Kamondi, A., Acsady, L. & Buzsaki, G. Eur. J. Neurosci. 10,
turn could activate kinases, such as myosin light-chain kinase
(MLCK), controlling vesicle recycling and the mobilization of a
6. Kamiya, H. & Zucker, R. S. Nature 371, 603–606 (1994).
vesicle reserve pool12. Ca2+ entry through L-type Ca2+ channels
7. Jensen, K., Jensen, M. S. & Lambert, J. D. J. Physiol. (Lond.) 519, 71–84
may thus mobilize vesicles into the releasable pool to ensure steady
8. Jensen, K., Jensen, M. S. & Lambert, J. D. J. Neurophysiol. 81, 1225–1230
transmission following high-frequency bursts. Accordingly, we
observed a transient post-tetanic depression when L-type channels
9. Atluri, P. P. & Regehr, W. G. J. Neurosci. 18, 8214–8227 (1998). 10. Holt, E. H., Broadus, A. E. & Brines, M. L. J. Biol. Chem. 271, 28105–28111
were blocked, resembling the long-lasting (>20 s) recovery from
synaptic depression induced by extensive presynaptic trains13. With
11. Zuhlke, R. D., Pitt, G. S., Deisseroth, K., Tsien, R. W. & Reuter, H. Nature 399,
the potentiating L-type Ca2+ channel-dependent mechanism
12. Ryan, T. A. J. Neurosci. 19, 1317–1323 (1999).
engaged, the terminals would be protected against synaptic depres-
13. Kraushaar, U. & Jonas, P. J. Neurosci. 20, 5594–5607 (2000).
sion by the increase of the releasable pool. In vivo, GABAergic bas-
14. Bartos, M., Vida, I., Frotscher, M., Geiger, J. R. & Jonas, P. J. Neurosci. 21,
ket cells14 fire at frequencies in the 40 Hz (gamma) range5. Thus,
15. Whittington, M. A., Traub, R. D., Kopell, N., Ermentrout, B. & Buhl, E. H. Int.
it is likely that hippocampal gamma activity activates the L-type
J. Psychophysiol. 38, 315–336 (2000). nature neuroscience • volume 4 no 10 • october 2001
P A T I E N T M E D I C A L H I S T O R Y Date ______________________________________________Refer ing Physician __________________________________________ Primary Care Physician _______________________________________Patient’s Name ___________________________________________________________ Date of Birth ______________________ Age ______Reason for today’s visit and prior treatments _______
Dr Johan Conradie Western Diagnostic Pathology, Perth, Western Australia A GP requested a home visit for some “follow up 1. What pre-analytical factors may alter the bloods” on a 68 year-old female patient being treated for a urinary tract infection. Urea, electrolytes and 2. What clinical factors may cause raised creatinine were requested with clinical notes stating ‘Foll

 2001 Nature Publishing Group http://neurosci.nature.com
2001 Nature Publishing Group http://neurosci.nature.com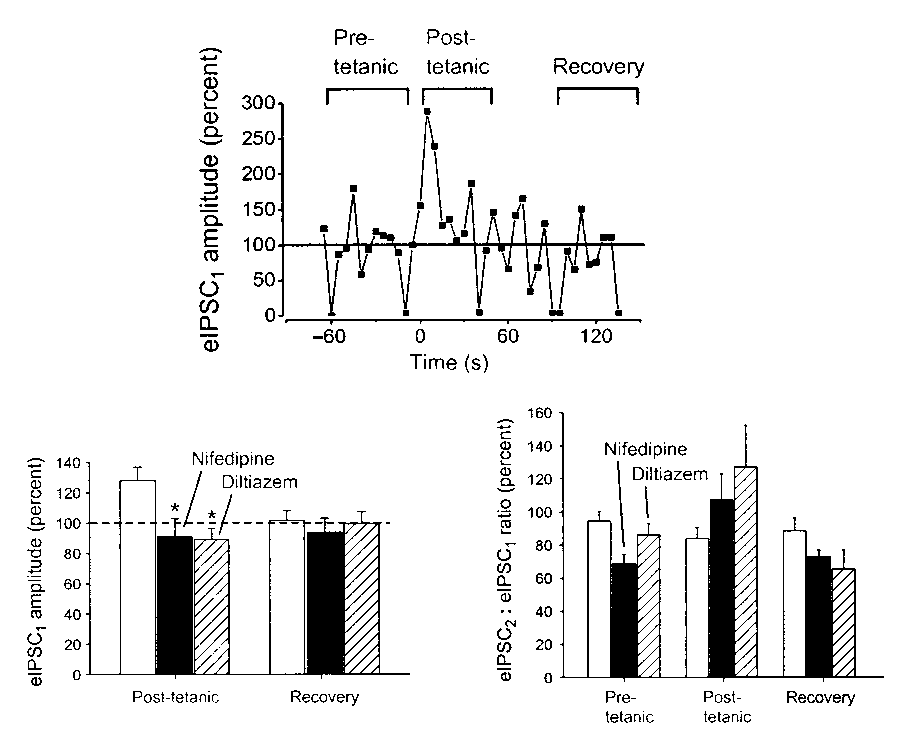

 2001 Nature Publishing Group http://neurosci.nature.com
2001 Nature Publishing Group http://neurosci.nature.com