Arch. Environ. Contam. Toxicol. 48, 242–250 (2005)DOI: 10.1007/s00244-003-0262-7
Comparative Sublethal Toxicity of Nine Pesticides on Olfactory LearningPerformances of the Honeybee Apis mellifera
A. Decourtye,1 J. Devillers,2 E. Genecque,3 K. Le Menach,4 H. Budzinski,4 S. Cluzeau,1 M. H. Pham-Delgue3
1 Association de Coordination Technique Agricole, Maison des Agriculteurs, La Tour de Salvagny, France2 CTIS, Rillieux La Pape, France3 Laboratoire de Neurobiologie ComparØe des InvertØbrØs, INRA, Bures-sur-Yvette, France4 Laboratoire de Physico-Toxico-Chimie des Systmes Naturels, UMR 5472 CNRS, UniversitØ de Bordeaux I, Talence Cedex, France
Received: 4 January 2004 /Accepted: 23 June 2004
Abstract. Using a conditioned proboscis extension response
(Waller et al. 1984; Bendahou et al. 1999; Decourtye et al.
(PER) assay, honeybees (Apis mellifera L.) can be trainedto
2004a), such effects also couldbe usedto better estimate the
associate an odor stimulus with a sucrose reward. Previous
hazardof pesticides to bees. Moreover, it is noteworthy that
studies have shown that observations of conditioned PER were
the EPPO guidelines require recording all abnormal behavioral
of interest for assessing the behavioral effects of pesticides on
effects observedduring the experiments (EPPO 1992).
the honeybee. In the present study, the effects of sublethal
Semifieldtests, representing more realistic exposure con-
concentrations of nine pesticides on learning performances of
ditions than in laboratory, have been cited as providing good
worker bees subjectedto the PER assay were estimatedand
information for the behavioral toxicity assessment of pesti-
compared. Pesticides were tested at three concentrations. The
cides (Cluzeau 2002). However, the regulatory guidelines give
highest concentration of each pesticide corresponded to the
only very limitedinformation on the type of behavioral data
median lethal dose value (48-h oral LD50), received per bee
that should be collected during the studies or how they should
and per day, divided by 20. Reduced learning performances
be included and interpreted in the risk assessment scheme
were observedfor bees surviving treatment with fipronil,
(Thompson andBrobyn 2002). Moreover, the semifieldtests,
deltamethrin, endosulfan, and prochloraz. A lack of behavioral
even if they are well suited, are technically difficult to main-
effects after treatment with k-cyalothrin, cypermethrin, s-flu-
valinate, triazamate, and dimethoate was recorded. No-ob-
necessity to have trainedpeople to carry them out are bounds
served-effect concentrations (NOECs) for the conditioned PER
limiting the number of facilities able to perform them in
were derived for the studied pesticides. Our study shows that
practice. Thus, the identification of precise behavioral effects
the PER assay can be usedfor estimating sublethal effects of
requires additional and specific methods to make appropriate
pesticides on bees. Furthermore, comparisons of sensitivity as
hazardassessment (Pham-Delgue et al. 2002). Consequently,
well as the estimation of NOECs, useful for regulatory pur-
the conditioned proboscis extension response (PER) assay
shouldbe use to overcome these problems (Decourtye andPham-Delgue 2002).
The PER assay tentatively reproduces what happens in
honeybee–plant interaction: when landing on the flower, theforager extends its proboscis as a reflex when the gustatory
The hazardassessment of pesticide toxicity to honeybees (Apis
receptors set on the tarsae, antennae, or mouth parts are
mellifera L.) is commonly estimatedfrom laboratory studies
stimulatedwith nectar. This reflex leads to the uptake of nectar
(median lethal dose: LD50) and from semifield and field
and induces the memorization of the floral odors diffusing
experimentations when the pesticides demonstrate a hazard
concomitantly. Once memorized, the odors play a prominent
quotient (application rate/LD50) over 50, or when they have a
role in flower recognition during the next trips (Menzel et al.
specific mode of action (e.g., insect growth regulators), or
1993). Consequently, an individual associative learning pro-
when there are indications of indirect effects such as delayed
cess is important for the effective accomplishment of foraging
action (EPPO 1992). Because behavioral effects of pesticides
activities. The associative learning of workers, investigated
in the honeybee have been shown to have the potential to
with the PER assay, may therefore be regarded as having a
induce a significant impact on the development of colonies
high ecological significance because it is a prerequisite to theforaging success of the whole colony.
The PER has been successfully reproduced under artificial
conditions (Kuwabara 1957; Takeda 1961), and has become a
Correspondence to: A. Decourtye; email: axel.decourtye@acta.asso.fr
valuable tool in studying various aspects of olfactory learning
Toxicity of Pesticides on Olfactory Learning of Honeybee
Table 1. Concentrations of agricultural chemicals applied with subchronic exposure before the conditioning procedure
a Actual concentrations of deltamethrin equal to 960, 429, and 212 lg Æ L)1. b Actual concentrations of cypermethrin equal to 782, 388, and207 lg Æ L)1.
processes (Bitterman et al. 1983; Menzel et al. 1993; Sandoz
To confirm the usefulness of the PER assay as a behavioral
et al. 1995). The PER assay with restrainedworkers has also
toxicity assessment method, the goal of our study was to
been usedto investigate the behavioral effects of pesticides
compare the effects of sublethal exposure of nine pesticides on
(Taylor et al. 1987; MamoodandWaller 1990; Stone et al.
the olfactory learning performances of worker bees subjected
1997; Abramson et al. 1999; Abramson andBoyd2001;
Weick andThorn 2002; Decourtye et al. 2003; Abramsonet al. 2004).
A previous work studying the behavioral toxicity of imi-
dacloprid and deltamethrin to bees indicated that a goodrelationship was foundbetween effects on olfactory responsesin free-flying foragers and in individuals subjected to the PER
paradigm (Decourtye et al. 2004a). The controlledconditions,the relationship with fieldcond
The nine studied pesticides (Table 1) were all technical grade. Del-
quantify the behavior pattern numerically ledus to assume that
tamethrin andprochloraz were obtainedfrom Hoechst Schering Ag-
the use of the PER assay, as a methodto evaluate the potential
rEvo S.A. (Aventis CropScience, France). All the other compounds
effect of pesticides on the honeybees foraging behavior, could
were purchasedfrom Cluzeau Info Labo (Sainte-Foy-La-Grand
help us to assess the toxicity of pesticides in a more compre-
France). Their purity was at least 98%, except s-fluvalinate, which
hensive way than by only considering lethality as currently
made in practice (Devillers 2002). However, a survey of the
The pesticides were tested at three different concentrations, with a
geometrical progression of factor 2. The highest testedconcentration
literature showedthat only a limitednumber of chemicals had
corresponded to the median lethal dose value (LD50 determined 48 h
been tested, and the studies using the PER assay were usually
after the oral treatments) divided by 20 (Table 1). From previous
not directly comparable because they were based on different
results (Decourtye et al. 2003), it was assumedthat this ratio belonged
methods for the administration of chemicals, the behavioral
to a sublethal domain. The 48-h LD50s reported in Table 1 were
response, andso on. Moreover, in these works only one dose,
previously determined from acute oral toxicity tests for deltamethrin,
not necessarily sublethal, was generally tested.
k-cyalothrin, andfipronil (Decourtye 2002), andfrom information
gainedin the existing literature for the other chemicals. The con-
was elicitedafter 3 s by contacting the antennae with a sucrose
centrations were calculatedfor a consumption of syrup estimatedto
solution (300 g Æ L)1) as the unconditioned stimulus, and the same
33 ll/bee/day (Decourtye et al. 2003).
solution was immediately given as a reward, before the odor delivery
Stock solutions with a given concentration of each chemical were
ended. Three successive conditioning trials (Cond1–Cond3) were
preparedin acetone (Table 1). Acetone was chosen following the
carriedout, followedby five test trials (Test1–Test5). The time
EPPO guidelines, because it is a rather generalist solvent (EPPO
interval between trials was 20–30 min. Conveniently, the positive
1992). Aliquots of the stock solutions were usedto make each test
responses at T1 of the individuals are scaled to 100 in order to better
solution at a specific concentration. The chemicals were added to a
characterize the extinction slope. During a test trial, the conditioned
500 g L-1sucrose solution. The final concentration of acetone in the
stimulus (pure linalool) was delivered for 6 s. The conditioned PER
sucrose solutions was 1% (vol/vol). The effects of insecticide-added
was recorded as a yes-or-no response (i.e., 0 or 1) when the odor alone
solutions were comparedwith that of an untreatedsucrose solution
was delivered during the 6 s of the test trial.
(with 1% acetone vol/vol). Fresh dosing solutions were prepared foreach test.
Samples of contaminatedsucrose solutions of d
cypermethrin delivered to bees were analyzed by gas chromatography/
mass spectrometry (K. Le Menach and H. Budzinski, unpublished).
For each chemical, the mortality accumulatedover 11 days of expo-sure was comparedbetween each concentration andthe control bymultiple two-by-two v2 tests with 1 df. To ensure that the experiment
error rate was a = 0.05, each comparison was carriedout according tothe Dunn-Sidak method (Sokal and Rohlf 1995) at a critical proba-bility of a' = 1 – (1 – a)1/k, where k was the number of intended tests.
Experiments were carriedout with worker bees of Apis mellifera
The significance level was 0.0085 for two-by-two comparisons of the
ligustica L. They were conducted with bees collected from outdoor
responses to three concentrations of each chemical andone control
hives. Emerging worker bees were caged in groups of 60 individuals.
They were provided with sugar food (mixture of sugar and honey),andwater ad libitum during the 2 first days and with pollen for the
The number of initial reflex responses andthe number of condi-
tionedresponses in each trial were comparedbetween the three
next 8 days. After 2 days, bees were continuously fed with sucrose
concentrations of each chemical andthe control by multiple two-by-
solution contaminated or not during 11 consecutive days. The feederswere changeddaily with fresh sucrose solutions. The bees were kept
two v2 tests with 1 df, with a critical probability level of 0.0085,according to the Dunn-Sidak correction of the standard probability
in an incubator (33 € 2°C, 40 € 10% relative humidity, darkness)
level. When conditions of application of the v2 test were not fulfilled
until 14–15 days old, and were used in the PER assay. It has been
according to the Cochran's rule, the Fisher's exact method was applied
shown that on average, worker bees become foragers at that age(Sakagami 1953; Seeley 1982) andgive the most consistent perfor-
mances in the conditioned proboscis extension assay (Pham-Delgueet al. 1990).
For bees from 2 to 14–15 days old, the quantity of the contaminated
During the treatment period(i.e., 11 days) for the nine tested
sugar solution provided daily was adjusted to the number of survivors.
pesticides, the volumes of syrup consumed for control (from
The mortality and consumption of syrup were recorded daily, and
22.0 to 45.2 ll/bee/day) and pesticide-treated groups (from
dead bees were discarded. Every testing day was organized as follows:
23.6 to 44.7 ll/bee/day) are not significantly different
bees previously exposedto three concentrations of each chemical
(ANOVA, 3 df, P > 0.05, in all cases). The geometrical pro-
were tested, as well as untreated control bees, leading to a total of 60–
gression of factor 2 between the different concentrations of
80 bees testedper day, with 16–20 bees for each treatment. Experi-ments were replicatedat least three times, until about 50–60 bees per
chemicals was respectedon the whole. These results suggest
that the tested concentrations for all pesticides do not have
After treatment, the bees were mounted individually in glass tubes
antifeedant effect on honeybees. The volumes of syrup con-
with only their antennae andmouth parts left free. They were starved
sumedare in agreement with the consumption initially esti-
for 4 h prior to conditioning. They were selected for showing a pro-
mated(33 ll/bee/day; Decourtye et al. 2003). Consequently,
boscis extension reflex after stimulation of the antennae with a su-
the quantities of chemicals actually ingestedby bees are close
crose solution (300 g Æ L)1). The number of individuals exhibiting the
reflex response was recorded. The ability to produce the reflex re-sponse reflects the state of the sensory-motor pathway underlying thePER. The general stimulation conditions as well as the conditioningand testing procedures were adapted from the work of Bitterman et al.
(1983) and are detailed in Sandoz et al. (1995). Bees were then placedin an airflow (main airflow of 50 ml Æ s)1 added to a secondary airflow
Cumulative mortality in bees significantly increases from that
of 2.5 ml Æ s)1) for 15 s, to be familiarizedwith the mechanical
of the control groups only with dimethoate and fipronil (Ta-
stimulation and with the experimental background. For the condi-tioning trials, the conditioned stimulus (10 ll of pure linalool, a
ble 2). A significant increase in mortality occurs with
standard floral odor, soaked on a filter paper strip inserted in a Pasteur
dimethoate at concentration of 580 lg Æ L)1 (28% versus 9.3%
pipette cartridge; Sigma, 95–97% purity) was delivered through the
mortality after 11 days, in the treated and control groups,
secondary flow (2.5 ml Æ s)1) for 6 s. During odor delivery, the PER
respectively; v2 = 9.3, 1 df, P = 0.002). The number of dead
Toxicity of Pesticides on Olfactory Learning of Honeybee
Table 2. Effects of subchronic exposures of nine agricultural chem-
bees in the groups exposedto fipronil at concentrations
icals on the survey andreflex responses of the honeybees
ranging from 2.2 to 9 lg Æ L)1 (40.6–91.1% mortality) aresignificantly different (v2, 1 df, P < 0.0083, in both cases) from
that of the control group (6.6% mortality). Consequently,
feeding honeybees with the sucrose solutions with added del-
tamethrin, prochloraz, endosulfan, k-cyalothrin, cypermethrin,
s-fluvalinate, or triazamate might be considered as sublethal,
contrary to fipronil anddimethoate treatments, which are le-
The comparison of the number of reflex responses obtained
when the antennae were contactedwith a sucrose solution, in
treatedandcontrol bees, was usedto evaluate the effects of the
pesticides on the gustatory and motor functions of the PER. At
least 66% of bees show a clear PER. For all chemicals, the
same level of reflex response in treatedanduntreatedbees is
found(v2, 1 df, P > 0.0083, in all cases; Table 2). This sug-
gests that the exposure to pesticides tested does not disrupt the
sensory andmotor components controlling the PER.
Table 3 shows the olfactory learning performances represented
as the percentage of conditioned PER obtained during the
training (Cond1–Cond3) and testing (Test1–Test5) phases, in
bees feeding the three concentrations of each pesticide and in
the control bees feeding only sucrose. Different letters indicate
significantly different response levels (v2 test or FisherÕs exact
method, 1 df, P < 0.0083). The results for deltamethrin are
provided in Figure 1, as an illustrative example of the learning
curves that can be drawn in the PER assay.
The percentage of bees treatedwith the highest d
dimethoate (580 lg Æ L)1) extending their proboscis in response
to the first presentation of odor (spontaneous responses ob-
servedat Cond1) is significantly higher than is observedwith
untreatedbees (36% versus 6%; v2 = 7.8, 1 df, P = 0.0052).
The feeding of workers with sucrose solution contaminated
with deltamethrin, prochloraz, endosulfan, or fipronil induces
significantly lower responses comparedto the untreatedbees,
considering Cond2–Cond3 for deltamethrin and Test4 for the
others pesticides. A reduction of the olfactory learning per-
formances is noted during conditioning trials in bees treated
with the highest concentration of deltamethrin (nominal and
actual concentrations of 940 and960 lg Æ L)1, respectively):
48% and60% conditioned responses at Cond2 and Cond3,
respectively, versus 60% and84% in the control (v2, 1 df, P <
0.0083, in both cases). At the testing trial Test4, lower levels
of responses are obtainedwith the highest dose of: prochloraz
(150 mg Æ L)1), reaching 36% of conditioned responses versus
N, number of bees per treatment group.
73% in the control group (v2 = 8.2, 1 df, P = 0.0048); endo-
For each chemical, the number of the cumulatedmortality in treated
sulfan (8 mg Æ L)1), reaching 6% of conditioned responses,
groups andin the control group were comparedusing v2 test or
versus 45% in the control group (v2 = 7.8, 1 df, P = 0.0037);
FisherÕs exact methodwith 1 df (P < 0.0083). Different letters indicatesignificantly different response levels.
fipronil (4.5 lg Æ L)1), reaching 7% of conditioned responses,versus 56% in the control group (v2 = 12.5, 1 df, P < 0.001). Conversely, for the above pesticides, no behavioral effect isobservedin the last training trial (Test5) (Table 3).
Table 3. Effects of subchronic exposures of nine agricultural chemicals on the learning performances of the honeybee
a N, number of bees per treatment group. b For each chemical, the number of the conditioned responses in treated groups and those in the control group were compared using v2 test orFisherÕs exact methodwith 1 df (P < 0.0083). Different letters indicate significantly different response levels. c Positive responses at T1 are scaledto 100.
Toxicity of Pesticides on Olfactory Learning of Honeybee
Fig. 1. Learning performances of deltamethrin-treated bees during conditioning (A) and testing (B) procedures of PER assay
In all trials, the level of responses of bees exposedto
The originality of our approach consists in taking into ac-
k-cyalothrin, cypermethrin, s-fluvalinate, andtriazamate is
count different concentrations in the PER assays. The deter-
equivalent to that obtainedwith control bees (v2, 1 df, P >
mination of the thresholdtoxicity concentrations is also
0.0083, in all cases). For these four chemicals, 66–93% of
possible. Thus, the no-observed-effect concentration (NOEC)
conditioned responses are obtained in treated bees at the last
for the conditioned PER is set to 2.2 lg Æ L)1, 470 lg Æ L)1
conditioning trial (Cond3) and 71–83% in the untreatedbees.
(actual concentration equals 429 lg Æ L)1), 4 mg Æ L)1, and75 mg Æ L)1 for fipronil, deltamethrin, endosulfan, and pro-chloraz, respectively. Considering the consumption of con-taminatedsyrup andthe number of bees, we can estimate that
the no-observed-effect dose of pesticide received per bee andper day is 0.07 ng for fipronil (LD50 divided by 80), 15 ng for
It is of interest to characterize honeybee behaviors that can be
deltamethrin (LD50 divided by 40), 125 ng for endosulfan
routinely used as indicators of sublethal exposure to pesticides.
(LD50 divided by 40), and 2.5 lg for prochloraz (LD50 di-
The possible long-term exposure to a toxic agent by contam-
vided by 40). Thus, fipronil is the most effective of the above
ination of storedfoodhas been establishedby studying the
pesticides tested to induce learning performances impairment.
transfer into the colony of pesticides sprayed on a crop (Fries
Under similar experimental conditions, the NOECs for imi-
andWibran 1987; Koch andWeisser 1997; Russel et al. 1998;
dacloprid and hydroxy-imidacloprid were estimated to 6 and
Villa et al. 2000). Thus, it is necessary to evaluate the viability
60 lg Æ L)1, corresponding to the DL50 value divided by 160
of worker bees newly involvedin foraging duties basedon
and80, respectively (Decourtye et al. 2003). As regards k-
their learning ability, after being fedwith a contaminatedfood
cyalothrin, cypermethrin, s-fluvalinate, andtriazamate, we can
within the hive. The preconditioning treatment applied in the
only say that the NOECs of these pesticides are superior to
present study leads to determining whether or not a pesticide
360 lg Æ L)1, 690 lg Æ L)1 (actual concentration of 782 lg Æ
exposure appliedprior to a learning task may affect the bees'
L)1), 300 mg Æ L)1, and600 lg Æ L)1, respectively.
performances. Among the nine pesticides tested, only fipronil,
To evaluate the usefulness of PER as a measure for toxicity
deltamethrin, endosulfan, and prochloraz yielded behavioral
assessment, it is necessary to compare these responses to
effects during the PER assay. This is consistent with previous
standard toxicity endpoints such as mortality. Learning per-
works reporting that the PER assay was adapted to the
formances after treatment with the highest concentration of
screening of the adverse effects of various pesticides to bees
deltamethrin, endosulfan, or prochloraz are decreased, in
(Taylor et al. 1987; MamoodandWaller 1990; Stone et al.
contrast to survival, which is not affected. The NOEC of hy-
1997; Abramson et al. 1999; Abramson andBoyd2001;
droxy-imidacloprid for the mortality was estimated to be
Weick andThorn 2002; Decourtye et al. 2003; Abramson et
120 lg Æ L)1, whilst the NOEC for the conditioned responses
al. 2004). Conversely, our results clearly indicate that the
was establishedat 60 lg Æ L)1 (Decourtye et al. 2003). From
range of testedconcentrations of k-cyalothrin, cypermethrin,
this study, it appears that most often the impairments in
s-fluvalinate, andtriazamate does not affect the learning per-
olfactory learning abilities are shown for chemical concen-
formances of bees. However, cypermethrin and s-fluvalinate
trations at which no additional mortality occurred.
are less toxic to honeybees than k-cyalothrin andtriazamate.
The choice of sublethal concentrations of pesticides is a
These results corroborate those of Taylor et al. (1987) showing
crucial problem when an attempt is made to estimate the ef-
that among a set of six pyrethroids, cypermethrin and s-flu-
fects of pesticides on bee behaviors. In this study, for each
valinate yielded the least impact on the honeybee learning.
chemical, the highest testeddose was the 48-h oral LD50 value
divided by 20. Considering the low mortality observed for
already established when the testing phase occurs. Delta-
most of the testedpesticides, it appears that this choice was
methrin wouldaffect the first step of information storage,
whereas endosulfan, fipronil, and prochloraz would interfere
In case of lethal treatment, the exposure to insecticide can
with the retrieval process resulting in the capacity to restore
result in a selection of worker bees staying alive because they
the conditioned response. However, further work is still nee-
are less sensitive to this pesticide than the other congeners.
ded to investigate more precisely the effects of these chemicals
Such tolerant bees can give an intact conditioned response
on the different parameters of the memory (acquisition, re-
level in the PER assay. For example, bees treatedwith DecisÒ
trieval, short-, medium- and long-term memory) during an
(0.5% a.i. deltamethrin) at a high dose exhibited similar pat-
olfactory conditioning of the PER, as investigated with imi-
tern of learning performances than control bees (Abramson
et al. 1999). In the current study, an adverse effect of
Besides behavioral effects of fipronil, an increase in the
dimethoate at its highest concentration (580 lg Æ L)1) is shown
mortality after 11 days appears in bees treated with this pes-
on survival of honeybees, but not on their learning perfor-
ticide. The lowest lethal dose of fipronil (0.1 ng per bee per
mances. Previous studies have assessed the effect of chlor-
day corresponding to a concentration of 2.2 lg Æ L)1) is 60
times lower than the LD50 value. At the same time, a lethal
dimethoate, on the behavior of parasitoids (Leptopilina het-
effect is significantly observedfor bees exposedto the highest
erotoma). Females of parasitoids were conditioned to associate
dose of dimethoate (20 ng per bee per day corresponding to a
an odor with the oviposition in host larvae of Drosophila
concentration of 580 lg Æ L)1). Although the long-term lethal
(Rafalimanana et al. 2002). Parasitoids exposed to the LD20
effect of dimethoate was previously demonstrated (Waller et
value of chlorpyrifos ovipositedthe host larvae more quickly
al. 1984), we have determined for the first time the chronic
than controls did. In our experiment, higher levels of sponta-
toxicity of fipronil to the honeybee. Using a similar laboratory
neous responses were obtainedin bees treatedwith the highest
chronic oral test with bees fedwith contaminatedsyrup,
concentration of dimethoate (580 lg Æ L)1). Thus, current re-
chronic toxicity can be foundeven at low concentrations of
sults andthose foundin the literature suggest that the high
imidacloprid (Suchail et al. 2000; Decourtye et al. 2003; De-
behavioral response levels in organophosphorus-treatedinsects
chaume-Moncharmont et al. 2003). In chronic toxicity studies,
were probably linkedto pharmacological action. These
imidacloprid reacts at doses 60 to 6000 times lower than those
chemicals act by inhibiting acetylcholinesterase andconse-
uce the same effect in acute intoxication
quently by prolonging activity of synapses (Padilla 1995). We
studies (Suchail et al. 2001). Thus, the acute toxicity tests,
assume that the increase in spontaneous responses in dimeth-
performed according to the EPPO guidelines (EPPO 1993),
appear to give only a partial measure of the lethal effects
perception or of response motricity. To confirm this hypoth-
because of the short duration of these tests (1 to 3 days in most
esis, further experiments shouldbe necessary. Works by
cases). Now, when the acute lethal effect is not obvious,
Abramson et al. (1999) shouldprovide some insight to per-
additional testing could give information on the long-term
lethal effects possibly induced by the toxic, as that was proved
The adverse effects are observed during a conditioning or
testing procedure according to the chemical tested. Ingestion
Although spraying of dimethoate-based formulations is
of deltamethrin significantly reduces the level of conditioned
prohibitedon flowering crops, the fieldapplication of formu-
responses in the conditioning procedure. This result suggests
lations containing deltamethrin, fipronil, or imidacloprid is
an adverse effect of deltamethrin on the ability of treated
allowed. In the current study, concentrations of 455 and
animals to learn the temporal relation between the uncondi-
227.5 lg Æ L)1 of deltamethrin (actual concentrations of 429
tioned stimulus and the conditioned one. In addition to con-
and212 lg Æ L)1) were tested. They are realistic because
ditioning procedure, the testing procedure points out the
500 lg Æ L)1 corresponds to the maximum concentration
resistance of bees to extinguish the response to a conditioned
measuredin oilseedrape flowers after spraying of De-
stimulus no longer associatedwith a reward. Abramson et al.
cisÒMicro (CETIOM unpublisheddata). We notedthe absence
of lethal andbehavioral effects after administration of these
recommended to control the cotton boll weevil influenced
concentrations. In an outdoor flight cage, representing more
extinction of the conditioned response. The authors suggested
realistic exposure conditions than those performed in a labo-
that motor system disruption was responsible for this event
ratory, a sugar solution containing 500 lg Æ L)1 of delta-
rather than an effect on the learning process. Our results
methrin offeredto a colony hadno effects on an olfactory
clearly indicate that endosulfan, as well as fipronil and pro-
learning discrimination task in free-flying foragers and in the
chloraz, do not affect either the reflex response or the condi-
PER procedure of restrained individuals (Decourtye et al.
tioned response level in the conditioning procedure, but the
2004a). Thus, the intact learning performances in treatedbees
decrease of response level in the testing procedure occurs more
at a realistic concentration of deltamethrin during a PER assay
rapidly compared to the control group.
are in agreement with those obtainedin semifieldconditions,
The conditioning and testing phases are two independent
at the colony level. This suggests that in fieldcond
foraging bees couldnot suffer from behavioral effects of
exposure. This may rely on the fact that different steps of the
deltamethrin after visiting of flowering crops treated with
memorization are involved. If we refer to the model of
DecisÒMicro. Contrary to the current study, the impact of
memory temporal schedule in the honeybee as described by
deltamethrin has been shown on survival of worker bees in the
Menzel (1999), the conditioning covers the information stor-
flight cage. This discrepancy might result in differences be-
age in the short-term memory, whilst long-term memory is
tween the dose received per bee and per day. In the flight cage
Toxicity of Pesticides on Olfactory Learning of Honeybee
study, we can estimate that the highest dose of deltamethrin
by the French Ministry of the Environment (MATE-01133-Evaluation
receivedper bee andper day was about equal to the LD50
et rØduction des risques lies à l'utilisation des pesticides).
value, while the value of 30 ng obtainedin the current labo-ratory study corresponds to the LD50 divided by 20.
Fipronil and imidacloprid, being the active ingredient of the
RegentÒ andGauchoÒ formulations, respectively, are author-izedas a sunflower seedcoating. In France, imidaclopridand
Abramson CI, Aquino IS, Ramalho FS, Price JM (1999) The effect of
fipronil were accusedof being a cause for the decline of sun-
insecticides on learning in the Africanized honey bee (Apis mel-
flower honey production. It is suspected that these products or
lifera L.). Arch Environ Contam Toxicol 37:529–535
theirs metabolites couldmigrate into nectar or pollen of treated
Abramson CI, BoydBJ (2001) An automatedapparatus for condi-
sunflowers and induce deleterious effects in foraging bees after
tioning proboscis extension in honey bees (Apis mellifera L.).
ingestion of contaminatedfood. Despite the fact that several
Abramson CI, Squire J, Sheridan A, Mulder PG Jr (2004) The effect
semifield and field tests indicated that seed dressing with imi-
of insecticides considered harmless to honey bees (Apis mellifera
dacloprid posed no risk during sunflower flowering (CurØ et al.
L.): Proboscis conditioning studies using the insect growth reg-
2000; Schmuck et al. 2001), a behavioral effect in the labora-
ulators ConfirmÒ2F (Tebufenozide) and DimilinÒ2L (Dif-
tory can be foundat a concentration potentially encounteredin
lubenzuron). Environ Entomol 33:378–388
plant tissues (Decourtye et al. 2003). From 1999 onwards, the
Barnavon M (1987) ExpØrimentation en laboratoire et en plein champ
French Ministry of Agriculture decided to suspend the regis-
du fluvalinate. Principes pour un insecticide. DØfense VØgØtaux
tration of the seedtreatment product GauchoÒ in sunflowers
according to the precautionary principle. As with imidacloprid,
Bendahou N, FlØchØ C, Bounias M (1999) Biological and biochemical
our study shows that an effect of fipronil can be observed on the
effects of chronic exposure to very low levels of dietary cyper-
learning performances of bees in the range of 2.2 to 4.5 lg Æ
methrin (Cymbush) on honeybee colonies (Hymenoptera: Api-dae). Ecotoxicol Environ Saf 44:147–153
L)1. Additional experiments are needed to establish the
Bitterman ME, Menzel R, Fietz A, Schäfer S (1983) Classical con-
thresholdconcentration of fipronil or of its metabolites from
ditioning proboscis extension in honeybees (Apis mellifera).
which the forager bees couldbe exposedandpossibly induces
drastic bee population losses, as observed by French beekeepers
Chalvet-Monfrey K (1996) Synergie entre la deltamØthrine et le
in colonies foraging on sunflowers treatedwith RegentÒ.
prochloraze chez l'abeille (Apis mellifera L.): Hypothses de
Our application of the PER assay for pesticide toxicity
mØcanismes dÕaction testØes par modØlisation. PhD Thesis, Uni-
assessment ledto characterization of effects on a behavioral
endpoint related to ability to associate an odor stimulus with
Cluzeau S (2002) Risk assessment of plant protection products on
sucrose reward. In general, a detailed basic knowledge on
honey bees. In: Devillers J, Pham-Delgue MH (eds) Honey bees:
behavior relevant for honeybee assays in ecotoxicology is still
estimating the environmental impact of chemicals. Taylor andFrancis, London, pp 42–55
relatively scarce, andthis is especially true for the influence of
CurØ G, Schmidt HW, Schmuck R (2000) Results of a comprehensive
principal general test variables on foraging behavior. The
field research programme with the systemic insecticide imida-
crucial problem in behavioral toxicology is the lack of stan-
cloprid(Gaucho). In: Belzunces LP, PØlissier C, Lewis GB (eds)
dardization for the tests. Therefore, the PER assay could be a
Hazards of pesticides to bees. INRA Edition, Avignon, pp 49–59
useful tool in the studies on behavioral effects of pesticides,
Dechaume-Moncharmont FX, Decourtye A, Hennequet C, Pham-
especially on foraging behavior, since it guarantees a good
Delgue MH, Pons O (2003) Statistical analysis of the honeybee
control of bee-rearing conditions and of exposure to chemicals.
survival after chronic exposure to insecticides. Environ Toxicol
During bee-rearing under laboratory conditions, the quality of
the food and the olfactory environment of the individuals must
Decourtye A (2002) Etude de lÕimpact de produits phytopharmaceu-
be strictly controlledbecause these factors can later influence
tiques sur la survie et lÕapprentissage associatif chez lÕabeilledomestique (Apis mellifera L.). PhD Thesis, University Paris XI,
pesticide sensitivity (Wahl and Ulm 1983) and learning per-
formances in the PER assay (Sandoz et al. 2000), respectively.
Decourtye A, Pham-Delgue MH (2002) The proboscis extension
Our study shows that the observations on conditioned PER
response: assessing the sublethal effects of pesticides on the
can be of interest in the assessment of behavioral effects of
honey bee. In: Devillers J, Pham-Delgue MH (eds) Honey bees:
pesticides to the honeybee, because this endpoint can be used
estimating the environmental impact of chemicals. Taylor and
to compare different pesticides by accounting for various
concentrations for increasing the accuracy of the results and
Decourtye A, Lacassie E, Pham-Delgue MH (2003) Learning per-
for deriving NOECs. In a previous study, we showed that the
formances of honeybees (Apis mellifera L.) are differentially af-
behavioral toxicity of imidacloprid observed in laboratory
fected by imidacloprid according to the season. Pest Manag Sci
conditions at the individual level (conditioned PER assay) was
Decourtye A, Devillers J, Cluzeau S, Charreton M, Pham-Delgue
consistent with results obtainedin semifieldexperiments at the
MH (2004a) Effects of imidacloprid and deltamethrin on asso-
colony level (Decourtye et al. 2004a). For the pesticides tested
ciative learning in honeybees under semi-field and laboratory
in the current study, further studies are still needed to establish
conditions. Ecotoxicol Environ Saf 57:410–419
Decourtye A, ArmengaudC, Renou M, Devillers J, Cluzeau S,
Gauthier M, Pham-Delgue MH (2004b) Imidacloprid impairsmemory andbrain metabolism in the honey bee (Apis melliferaL.). Pestic Biochem Physiol 78:83–92
We wish to thank M. Charreton andB. Roger for
Devillers J (2002) Acute toxicity of pesticides to honey bees. In:
technical assistance in bee rearing. This work benefitedfrom a grant
Devillers J, Pham-Delgue MH (eds) Honey bees: estimating the
environmental impact of chemicals. Taylor andFrancis, London,
Sandoz JC, Roger B, Pham-Delgue MH (1995) Olfactory learning
andmemory in the honeybee: comparison of different classical
European andMediterranean Plant Protection Organisation (EPPO)
conditioning procedures of the proboscis extension response. C R
(1992) Guideline on test methods for evaluating the side-effects
of plant protection products on honey bees. EPPO Bull 22:203–
Sandoz JC, Laloi D, Odoux JF, Pham-Delgue MH (2000) Olfactory
information transfer in the honeybee: comparedefficiency of
European andMediterranean Plant Protection Organisation (EPPO)
classical conditioning and early exposure. Anim Behav 59:1025–
(1993) Decision-making scheme for the environmental risk
assessment of plant protection products. Chapter 10: honeybees.
Schmuck R, Schçning R, Stork A, Schramel O (2001) Risk to hon-
eybees (Apis mellifera L. Hymenoptera) by an imidacloprid seed
Fries I, Wibran K (1987) Effects on honey-bee colonies following
dressing of sunflowers. Pest Manag Sci 57:225–238
application of the pyrethroids cypermethrin and PP 321 in flow-
Seeley TD (1982) Adaptative significance of the age polyethism
ering oilseedrape. Am Bee J 127:266–269
schedule in honeybee colonies. Behav Ecol Sociobiol 11:287–293
Gough HJ, MacIndoe EC (1994) The use of dimethoate as a reference
Sokal RR, Rohlf FJ (1995) Biometry: the principles of practice of
compoundin laboratory acute toxicity tests on honey bees (Apis
statistics in biological research. WH Freeman andCo., New York
mellifera L.) 1981–1992. J Apic Res 33:119–125
Stevenson JH (1978) The acute toxicity of unformulatedpesticides to
Koch H, Weiber P (1997) Exposure of honey bee during pesticide
worker honey bees (Apis mellifera L.). Plant Pathol 27:38–40
application under field conditions. Apidologie 28:439–447
Stone JC, Abramson CI, Price JM (1997) Task-dependent effects of
Kuwabara M (1957) Bildung des bedingten Reflexes von Pavlovs
dicofol (Kelthane) on learning in the honey bee (Apis mellifera).
Typus bei der Honigbiene, Apis mellifica. J Fac Sci Hokkaido
Suchail S, Guez D, Belzunces LP (2000) Characteristics of imida-
MamoodAN, Waller GD (1990) Recovery of learning responses by
clopridtoxicity in two Apis mellifera subspecies. Environ Toxicol
honeybees following a sublethal exposure to permethrin. Physiol
Suchail S, Guez D, Belzunces LP (2001) Discrepancy between acute
Menzel R, Greggers U, Hammer M (1993) Functional organization of
and chronic toxicity induced by imidacloprid and its metabolites
appetitive learning andmemory in a generalist pollinator, the
in Apis mellifera. Environ Toxicol Chem 20:2482–2486
honey bee. In: Papaj DR, Lewis AC (eds) Insect learning.
Takeda K (1961) Classical conditioned response in the honey bee.
Menzel R (1999) Memory dynamics in the honeybee. J Comp Physiol
Taylor KS, Waller GD, Crowder LA (1987) Impairment of classical
conditioned response of the honey bee (Apis mellifera L.) by
Miniggio C, Borneck R, Arnold G (1990) Etudes des effets à long
sublethal doses of synthetic pyrethroid insecticides. Apidologie
terme des pesticides chez lÔabeille Apis mellifera. ComitØ de
lÕEcologie et de la Gestion du Patrimoine Naturel, Ministre de
Thompson H, Brobyn T (2002) Sub-lethal effects in honeybees: their
significance anduse in pesticide risk assessment. 8th International
Padilla S (1995) The neurotoxicity of cholinesterase-inhibiting
Symposium Hazards of Pesticides to Bees, Bologna, September
insecticides: past and present evidence demonstrating persistent
Vaughan A (1986) Cypermethrin: honey bee acute contact andoral
Pham-Delgue MH, De Jong R, Masson C (1990) Effet de l'âge sur la
LD50. U.S. Environmental Protection Agency, Environmental
rØponse conditionnØe d'extension du proboscis chez l'abeille
domestique. C R Acad Sci Paris, Ser III 310:527–532
Villa S, Vighi M, Finizio A, Bolchi Serini G (2000) Risk assessment
Pham-Delgue MH, Decourtye A, Kaiser L, Devillers J (2002)
for honeybees from pesticide-exposed pollen. Ecotoxicology
Methods to assess the sublethal effects of pesticides on honey
Wahl O, Ulm K (1983) Influence of pollen feeding and physiological
Rafalimanana H, Kaiser L, Delpuech JM (2002) Stimulating effects of
condition on pesticide sensitivity of the honey bee Apis mellifera
the insecticide chlorpyrifos on host searching and infestation
efficacy of a parasitoidwasp. Pest Manag Sci 58:321–328
Waller GD, Erikson BJ, Harvey J, Martin JH (1984) Effects of
Russel D, Meyer R, Bukowski J (1998) Potential impact of micro-
dimethoate on honeybees (Hymenoptera: apidae) when applied to
flowering lemons. J Econ Entomol 77:70–74
Weick J, Thorn RS (2002) Effects of acute sublethal exposure to
Sakagami SF (1953) Untersuchungen über die Arbeitsteilung in einem
coumaphos or diazinon on acquisition and discrimination of odor
zwergfolk der Honigbiene. Beiträge zur Biologie des Bienen-
stimuli in the honey bee (Hymenoptera: Apidae). J Econ Entomol
volkes, Apis mellifera L. I. Jpn J Zool 11:117–185
Allegato 10 PERCORSO DIAGNOSTICO-TERAPEUTICO DELLA SINDROME DI RETT PREMESSA La sindrome di Rett (RTT) è una grave patologia neurologica, che colpisce prevalentemente le femmine, con un’incidenza variabile tra 1:10000 e 1:15000. La patologia è stata descritta per la prima volta nel 1966 da parte del medico austriaco Andreas Rett, e successivamente resa nota a tutto il mondo dal
POST-OPERATIVE INSTRUCTIONS SPINE SURGERY After your surgery there are several points we would like you to keep in mind. Most patients will be able to maintain a fairly normal level of activity following surgery. We do ask that you adhere to the activity restrictions described, as well as note some of the other care instructions. We value your health, well-being and comfort. If you do h
 Arch. Environ. Contam. Toxicol. 48, 242–250 (2005)DOI: 10.1007/s00244-003-0262-7
Comparative Sublethal Toxicity of Nine Pesticides on Olfactory LearningPerformances of the Honeybee Apis mellifera
A. Decourtye,1 J. Devillers,2 E. Genecque,3 K. Le Menach,4 H. Budzinski,4 S. Cluzeau,1 M. H. Pham-Delgue3
1 Association de Coordination Technique Agricole, Maison des Agriculteurs, La Tour de Salvagny, France2 CTIS, Rillieux La Pape, France3 Laboratoire de Neurobiologie ComparØe des InvertØbrØs, INRA, Bures-sur-Yvette, France4 Laboratoire de Physico-Toxico-Chimie des Systmes Naturels, UMR 5472 CNRS, UniversitØ de Bordeaux I, Talence Cedex, France
Received: 4 January 2004 /Accepted: 23 June 2004
Abstract. Using a conditioned proboscis extension response
(Waller et al. 1984; Bendahou et al. 1999; Decourtye et al.
Arch. Environ. Contam. Toxicol. 48, 242–250 (2005)DOI: 10.1007/s00244-003-0262-7
Comparative Sublethal Toxicity of Nine Pesticides on Olfactory LearningPerformances of the Honeybee Apis mellifera
A. Decourtye,1 J. Devillers,2 E. Genecque,3 K. Le Menach,4 H. Budzinski,4 S. Cluzeau,1 M. H. Pham-Delgue3
1 Association de Coordination Technique Agricole, Maison des Agriculteurs, La Tour de Salvagny, France2 CTIS, Rillieux La Pape, France3 Laboratoire de Neurobiologie ComparØe des InvertØbrØs, INRA, Bures-sur-Yvette, France4 Laboratoire de Physico-Toxico-Chimie des Systmes Naturels, UMR 5472 CNRS, UniversitØ de Bordeaux I, Talence Cedex, France
Received: 4 January 2004 /Accepted: 23 June 2004
Abstract. Using a conditioned proboscis extension response
(Waller et al. 1984; Bendahou et al. 1999; Decourtye et al.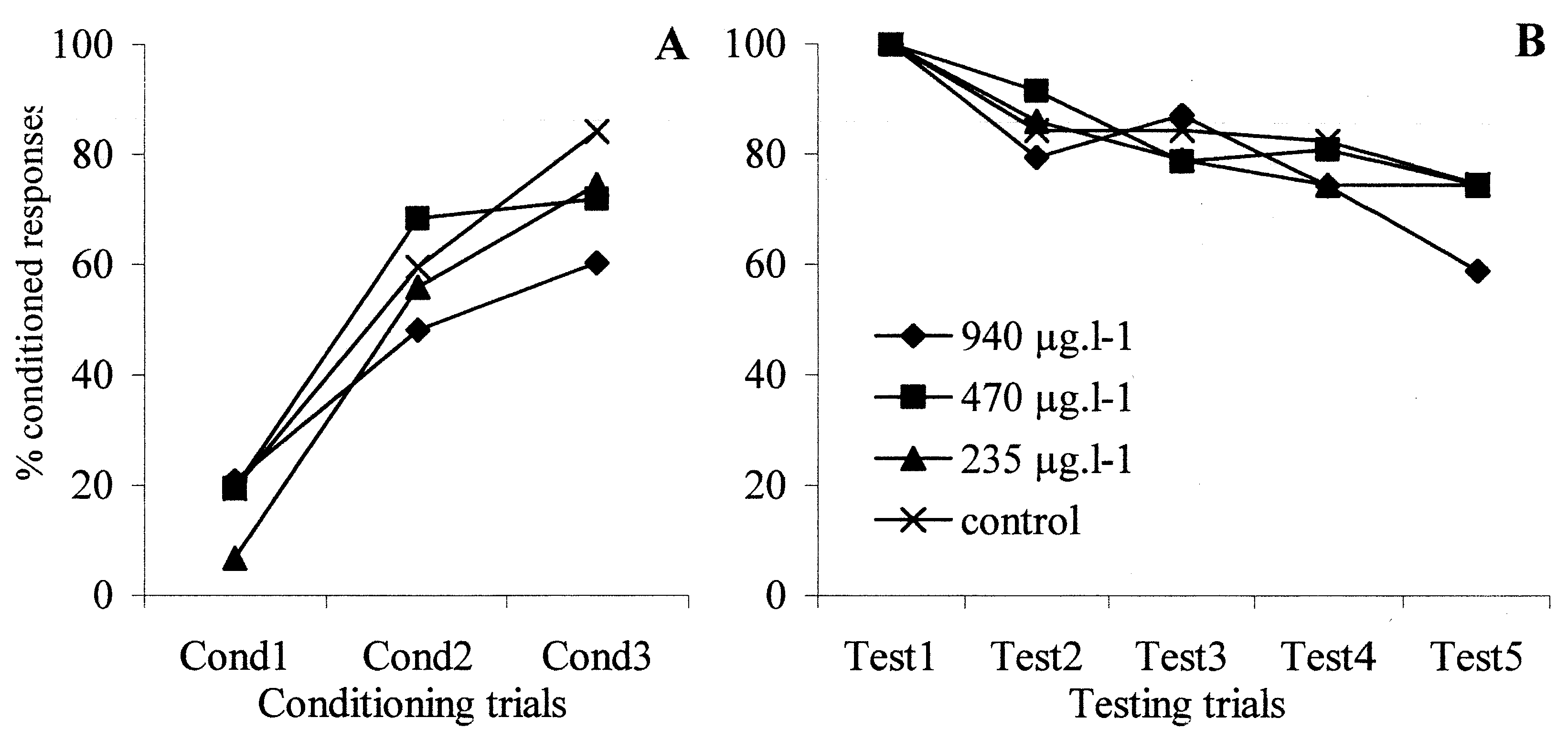 Toxicity of Pesticides on Olfactory Learning of Honeybee
Fig. 1. Learning performances of deltamethrin-treated bees during conditioning (A) and testing (B) procedures of PER assay
In all trials, the level of responses of bees exposedto
The originality of our approach consists in taking into ac-
k-cyalothrin, cypermethrin, s-fluvalinate, andtriazamate is
count different concentrations in the PER assays. The deter-
equivalent to that obtainedwith control bees (v2, 1 df, P >
mination of the thresholdtoxicity concentrations is also
0.0083, in all cases). For these four chemicals, 66–93% of
possible. Thus, the no-observed-effect concentration (NOEC)
conditioned responses are obtained in treated bees at the last
for the conditioned PER is set to 2.2 lg Æ L)1, 470 lg Æ L)1
conditioning trial (Cond3) and 71–83% in the untreatedbees.
Toxicity of Pesticides on Olfactory Learning of Honeybee
Fig. 1. Learning performances of deltamethrin-treated bees during conditioning (A) and testing (B) procedures of PER assay
In all trials, the level of responses of bees exposedto
The originality of our approach consists in taking into ac-
k-cyalothrin, cypermethrin, s-fluvalinate, andtriazamate is
count different concentrations in the PER assays. The deter-
equivalent to that obtainedwith control bees (v2, 1 df, P >
mination of the thresholdtoxicity concentrations is also
0.0083, in all cases). For these four chemicals, 66–93% of
possible. Thus, the no-observed-effect concentration (NOEC)
conditioned responses are obtained in treated bees at the last
for the conditioned PER is set to 2.2 lg Æ L)1, 470 lg Æ L)1
conditioning trial (Cond3) and 71–83% in the untreatedbees.