RAPID COMMUNICATION
GABA and Trk Receptor Signaling Mediates Long-Lasting
VIBHAKAR C. KOTAK,1 CHRISTOPHER DIMATTINA,1 AND DAN H. SANES1,21Center for Neural Science and 2Department of Biology, New York University, New York, New York 10003
Received 27 November 2000; accepted in final form 23 March 2001
Kotak, Vibhakar C., Christopher DiMattina, and Dan H. Sanes.
Stimulation of MNTB afferents at a low rate leads to a
GABA and Trk receptor signaling mediates long-lasting inhibitory
long-lasting depression of synaptic inhibition in LSO neurons
synaptic depression. J Neurophysiol 86: 536 –540, 2001. In many
(Kotak and Sanes 2000). This form of inhibitory synaptic
areas of the nervous system, excitatory and inhibitory synapses are
plasticity declines with age, and we have hypothesized that it
reconfigured during early development. We have previously described
contributes to the activity-dependent reorganization of MNTB
the anatomical refinement of an inhibitory projection from the medial
arbors within the LSO (Sanes and Taka´cs 1993). Although
nucleus of the trapezoid body to the lateral superior olive in thedeveloping gerbil auditory brain stem. Furthermore, these inhibitory
long-term inhibitory synaptic depression has been reported in
synapses display an age-dependent form of long-lasting depression
this and other systems (Komatsu 1994; Morishita and Sastry
when activated at a low rate, suggesting that this process could
1991; Oda et al. 1998), the signaling pathway that initiates this
support inhibitory synaptic refinement. Since the inhibitory synapses
form of plasticity has not been examined. In contrast, co-
release both glycine and GABA during maturation, we tested whether
activation of glutamatergic and GABAergic afferents can pro-
receptor signaling could initiate the decrease in synaptic
duce inhibitory depression through an N-methyl-D-aspartate
strength. When whole cell recordings were made from lateral superior
(NMDA) receptor mechanism (Caillard et al. 1999).
olive neurons in a brain slice preparation, the long-lasting depression
This present study focuses on two candidate signaling sys-
of medial nucleus of the trapezoid body– evoked inhibitory potentials
tems. First, the MNTB-evoked inhibitory response recorded in
the gerbil LSO is predominantly GABAergic before hearing
addition, inhibitory potentials could be depressed by repeated expo-sure to the GABA receptor agonist, baclofen. Since GABA receptor
onset and switches to a predominantly glycinergic input post-
signaling may not account entirely for inhibitory synaptic depression,
natally (Kotak et al. 1998). This finding suggested that
we examined the influence of neurotrophin signaling pathways lo-
GABAergic transmission could play a significant role during
cated in the developing superior olive. Bath application of brain-
inhibitory synaptogenesis. Second, MNTB neurons express
derived neurotrophic factor or neurotrophin-3 depressed evoked in-
neurotrophins, and LSO neurons express their cognate recep-
hibitory potentials, and use-dependent depression was blocked by the
tors during development (Hafidi 1999; Hafidi et al. 1996).
tyrosine kinase antagonist, K-252a. We suggest that early expression
Since neurotrophin/Trk signaling pathways have been shown
of GABAergic and neurotrophin signaling mediates inhibitory synap-
to modulate synaptic transmission (Kang and Schuman 1995;
tic plasticity, and this mechanism may support the anatomical refine-
Kim et al. 1994; Levine et al. 1998), they may be relevant to
the plasticity displayed by MNTB synapses. Therefore we havetested whether signals mediated by GABA and neurotrophin
receptors are involved in the long-lasting depression of inhib-
Although neuronal discharge can be quite low during early
development, spontaneous and evoked activity has a profound
impact on the selective loss or survival of synaptic contacts
Gerbils (Meriones unguiculatus) aged postnatal days 8 –12 (P8 –
(Sanes et al. 2000a). Manipulations of excitatory transmission
12) were used to make 300-M coronal brain slices through the LSO
can disrupt the normal elimination of motor axons onto muscle
and MNTB. The artificial cerebrospinal fluid (ACSF) contained (in
fibers, and prevent the refinement of excitatory connections in
mM) 125 NaCl, 4 KCl, 1.2 KH PO , 1.3 MgSO , 26 NaHCO3, 15
the CNS (Cline et al. 1987; Ichise et al. 2000; Kleinschmidt et
al. 1987; O’Brien et al. 1978; Scherer and Udin 1989; Simon
with 95% O -5% CO ). The ACSF was continuously superfused in
the recording chamber at 4 –5 ml per min at room temperature (22–
et al. 1992; Thompson et al. 1979). There is now evidence that
24°C). Whole cell current-clamp recordings were obtained from LSO
inhibitory terminals also become refined during development.
neurons (Warner PC-501A), and 200-s electrical pulses were deliv-
In the gerbil lateral superior olive (LSO), the inhibitory affer-
ered directly to the MNTB, as described previously (Kotak and Sanes
ent fibers from the medial nucleus of the trapezoid body
2000). The internal patch solution contained (in mM) 127.5 potassium
(MNTB) become restricted anatomically during postnatal de-
gluconate, 0.6 EGTA, 10 HEPES, 2 MgCl , 5 KCl, 2 ATP, and 0.3
GTP (pH 7.2). To block tyrosine kinase in the postsynaptic LSO
Address for reprint requests: D. H. Sanes, Center for Neural Science,
The costs of publication of this article were defrayed in part by the payment
4 Washington Place, New York University, New York, NY 10003 (E-mail:
of page charges. The article must therefore be hereby marked ‘‘advertisement’’
in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
0022-3077/01 $5.00 Copyright 2001 The American Physiological Society
RECEPTOR–MEDIATED INHIBITORY DEPRESSION
neuron, K-252a (200 nM) was added to the internal pipette solution. To examine inhibitory synaptic depression, MNTB-evoked maximumamplitude inhibitory postsynaptic potentials (IPSPs) were first ac-quired during a 15-min baseline period initially every minute for thefirst 5 min and then at the 10th and 15th min (Kotak and Sanes 2000). The MNTB was then activated with low-frequency stimulation (LFS:1 Hz for 15 min). Immediately following LFS, MNTB-evoked IPSPswere recorded every min for the first 5 min and every 5 min thereafter. To block GABA
receptors, SCH-50911 (5–10 M, Tocris) was
bath-applied throughout the experiment beginning 5 min before re-cording the first IPSP.
In a separate set of experiments, IPSPs were recorded for about 1 h
at a very low rate of acquisition that does not produce synapticdepression (0.03 Hz), and the slices were exposed to either a GABABreceptor agonist (baclofen, 100 M, Sigma Chemicals), or a neuro-trophin [brain derived neurotrophic factor (BDNF), 50 –100 ng/ml,Sigma Chemicals or Alamone Laboratories; NT-3, 25–50 ng/ml,Sigma Chemicals]. In many of these experiments, contaminatingglutamatergic activity was blocked with 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX; 20 M) or kynurenic acid (4 mM). This was donefor control LFS experiments (n ϭ 3), baclofen exposure (n ϭ 2),BDNF exposure (n ϭ 7), NT-3 exposure (n ϭ 6), and SCH-50911treatment (n ϭ 3).
Data were collected using a Macintosh PPC running a custom-
designed IGOR (WaveMetrics, v3.14) macro called SLICE. The datawere analyzed off-line using a second IGOR macro called SLICEANALYSIS. Each macro is available with complete documenta-tion on-line at http://www.cns.nyu.edu/ϳsanes/slice_software. TheSLICE macro controls the stimulus isolation units and patch-clamp
Long-lasting depression of inhibitory transmission was mediated by
amplifier via an ITC-18 Computer Interface (Instrutech Corporation)
GABA receptors. A: medial nucleus of the trapezoid body (MNTB)– evoked
using an IGOR external operation commands (XOP version 2.6,
maximum inhibitory postsynaptic potentials (IPSPs) were recorded from thelateral superior olive (LSO) before and after low-frequency stimulation (LFS)
Instrutech). Data were sampled and stored at 10 kHz. Analyses of
of the MNTB. Example IPSPs are shown for a postnatal day 9 (P9) neuron
peak IPSP amplitude, rising slope, and duration were performed
recorded in control (top) or SCH-5091– containing ACSF (bottom). E
off-line. Data are presented as means Ϯ SE or as a percent of the
Ϫ53 and Ϫ52 mV, respectively. B: summary for all recorded LSO neurons at
normalized IPSP amplitudes as indicated in RESULTS and the figure
P8 –12 in the absence and presence of SCH-50911 (mean Ϯ SE). Synaptic
legends. All analyses were performed with the Student’s t-test.
depression was robust (43%) at 50 – 60 min following LFS when comparedwith pre-LFS IPSPs (●). Age-matched neurons treated with SCH-50911 (E)displayed a marginal change in IPSP amplitude following LFS. The mean
percent change was calculated by comparing the average normalized IPSPamplitude recorded at 50 – 60 min post LFS with the normalized mean initial
The data reported here are drawn from whole cell current-
IPSP amplitude (0%) during 1st 5 min of the recording session (for control
clamp recordings from 74 LSO neurons. Each recording was
neurons: t ϭ 5.1, df ϭ 18, P Ͻ 0.0001; for SCH-50911–treated neurons: t ϭ
Ϫ0.56, df ϭ 18, P ϭ 0.57).
obtained from a separate brain slice. In the initial experiments,MNTB-evoked maximum amplitude IPSPs were recorded
3-min intervals) induced a long-lasting depression. There was
without any pharmacological agents in the ACSF. As shown
also a significant decrease in the IPSP rising slope (50%
for a control P9 neuron in Fig. 1, the MNTB-evoked IPSP was
decline, P Ͻ 0.01). In three of four neurons tested, the LSO
about 11 mV during the pre-LFS period, but decreased to about
input resistance decreased by approximately 30% during ba-
6.5 mV following LFS treatment (top). The average IPSP
clofen exposure. In two additional experiments, a single dose
amplitude reduction was 43% at 1 h following LFS, as com-
exposure of baclofen (100 M) caused the MNTB-evoked
pared with the baseline IPSP amplitude prior to LFS (n ϭ 10).
IPSPs to decrease by about 50% for approximately 10 min.
In three recordings, ionotropic glutamate receptors were
This baclofen-elicited depression was eliminated when the
blocked with kynurenic acid (4 mM), and this did not alter the
slice was pretreated for 6 min with 10 M SCH-50911 (data
magnitude of depression (a 45% reduction in IPSP amplitude
not shown). This indicated that the synaptic- and agonist-
was observed). To assess the role of GABA receptors during
mediated depression involved the same receptor.
the initiation of inhibitory synaptic depression, we applied the
While the GABA receptor antagonist results suggest that
GABA receptor antagonist SCH-50911 (5–10 M) through-
this receptor is necessary for induction of inhibitory depres-
out the experiment, beginning 5 min before the first IPSP was
sion, additional mechanisms have not been ruled out. Therefore
recorded. As shown in Fig. 1, when LFS was delivered in the
two neurotrophin signaling systems (BDNF and NT-3) known
presence of SCH-50911, the magnitude of long-lasting depres-
to be localized to the MNTB-LSO pathway were tested as
sion was blocked as compared with the untreated controls.
candidates for a depression mechanism. For these experiments,
The second experimental strategy to assess GABA receptor
IPSPs were recorded every 30 s for approximately 1 h. In
involvement in inhibitory depression was an extension of our
control recordings, this stimulus rate did not alter IPSP ampli-
previous finding that baclofen reversibly depressed IPSPs fol-
tude significantly. The change in IPSPs was calculated by
lowing a single exposure (Kotak et al. 1998). As shown in Fig.
comparing the mean IPSP amplitude (ϮSE) recorded at 50 – 60
2, repeated perfusion (100 M baclofen; 5 ϫ 10 s exposures at
min with the mean initial IPSP amplitude (ϮSE) during first 10
V. C. KOTAK, C. DIMATTINA, AND D. H. SANES
by both auditory and visual experience (Knudsen and Brainard1991; Mogdans and Knudsen 1993). In the gerbil LSO, inter-aural level difference coding improves with age, and severalanatomical and physiological properties are disrupted by deaf-ferentation during development (Sanes et al. 2000b). We havepreviously shown that inhibitory projections from MNTB toLSO become refined during development, and this process isdisrupted by deafferentation (Sanes and Siverls 1991; Sanesand Taka´cs 1993). More recently, we have found that thestrength of these inhibitory synapses depends on activity, andthis phenomenon wanes with age (Kotak and Sanes 2000). Thepresent results suggest that use-dependent depression of inhib-itory synapses requires GABA receptors, and may also em-
Repeated activation of GABA receptors elicited long-lasting syn-
aptic depression. A: a maximum amplitude IPSP is shown for a P10 neuronbefore and after repeated baclofen exposure. B: the bar graph compares thepercent change in the normalized IPSP amplitude during a control period, a15-min drug exposure period, and after a recovery period. There was asignificant decline (asterisk) in IPSP amplitudes during baclofen treatment, andthis depression persisted at 30 min after the last baclofen exposure. The changein IPSPs was calculated by comparing the average normalized IPSP amplituderecorded during baclofen exposure and at 50 – 60 min of the experiments withthe initial IPSP amplitude (0%) during 1st 10 min of the recording session(comparison between initial IPSPs and IPSPs during baclofen exposure: t ϭ4.01, df ϭ 6, P Ͻ 0.007; comparison between initial IPSPs and IPSPs at 50 – 60min: t ϭ 4.08, df ϭ 6, P Ͻ 0.006).
min of the recording session (initial IPSP amplitude ϭ 8.7 Ϯ0.7 mV, mean Ϯ SE; IPSP amplitude at 50 – 60 min followingLFS ϭ 8.9 Ϯ 0.8 mV; t ϭ Ϫ0.51, df ϭ 10, P ϭ 0.620). Inseparate recordings, bath application of BDNF (50 –100 ng/ml)for 5– 8 min resulted in a small decrease in IPSP amplitude. Approximately 10 min after BDNF application, IPSP ampli-tudes declined, and this attenuation reached its maximum byabout 20 –30 min following drug exposure, but the change didnot reach significance (comparison between initial IPSPs andIPSPs during BDNF exposure: t ϭ 0.36, df ϭ 18, P ϭ 0.72;comparison between initial IPSPs and IPSPs at 50 – 60 min: t ϭ0.17, df ϭ 14, P ϭ 0.07). Exposure to NT-3 (25–50 ng/ml)produced a larger and more rapid decline in IPSP amplitude,and this decline was highly significant (Fig. 3). Finally, toassess whether neurotrophin receptors could influence synap-tically evoked depression, a tyrosine kinase antagonist (200nM K-252a) was added to the internal patch solution. Asshown in Fig. 3B, K-252a prevented LFS from inducing a
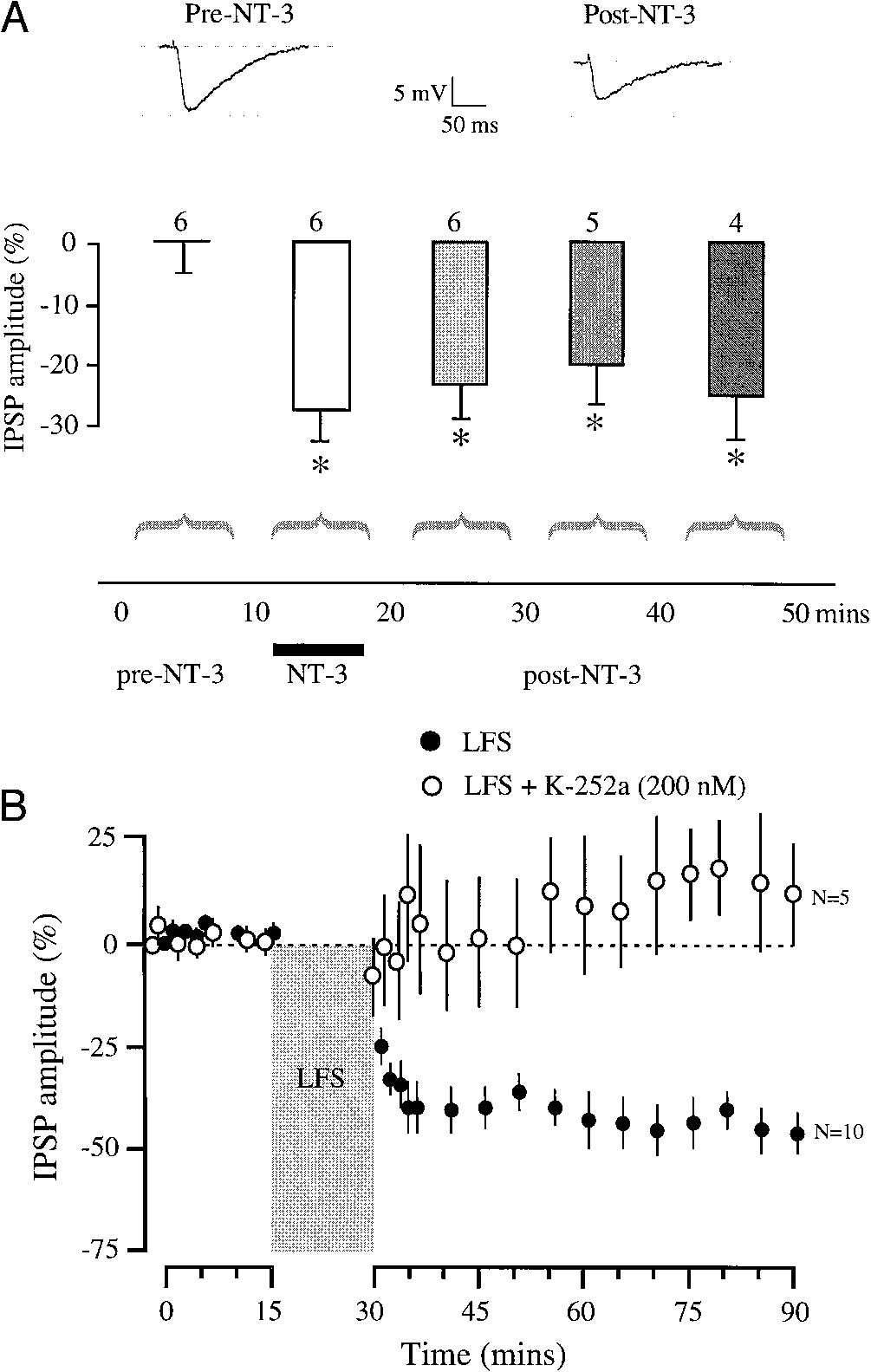
Neurotrophin signaling depresses inhibitory transmission. A: a
significant change in IPSP amplitude (mean initial IPSP am-
maximum amplitude IPSP is shown for a P11 neuron before and after NT-3(25 ng/ml) exposure for 8 min. The IPSP depressed by about 30%. The bar
plitude ϭ 9 Ϯ 1 mV; mean IPSP amplitude at 50 – 60 min
graph compares percent change in IPSP amplitude before, during, and after
NT-3 exposure. The IPSPs decreased significantly during NT-3 applicationwhen compared with pre-NT-3 treatment IPSPs (asterisk), and remained de-pressed at 40 –50 min (comparison between initial IPSPs and IPSPs during
NT-3 exposure: t ϭ 2.58, df ϭ 10, P ϭ 0.02; comparison between initial IPSPs
A number of studies suggest that auditory coding properties
and IPSPs at 50 – 60 min: t ϭ 4.65, df ϭ 8, P ϭ 0.001). B: summary for
mature postnatally, and that this improvement is due, in part, to
neurons in the absence and presence of K-252a (control data from Fig. 1). Neurons recorded with K-252a in the pipette solution (E) displayed no change
experience-dependent mechanisms (Sanes and Walsh 1997).
in IPSP amplitude following LFS (comparison between initial IPSPs and IPSPs
For example, sound localization in the barn owl is influenced
at 50 – 60 min: t-test; t ϭ Ϫ0.73, df ϭ 8, P ϭ 0.48).
RECEPTOR–MEDIATED INHIBITORY DEPRESSION
Inhibitory synapses in LSO are predominantly GABAergic
expression of long-term depression of GABAergic synapses in the neonatal
during the first two postnatal weeks, and gradually adopt a
rat hippocampus. J Neurosci 19: 7568 –7577, 1999.
CLINE HT, DEBSKI EA, AND CONSTANTINE-PATON M. N-methyl-D-aspartate
glycinergic phenotype (Kotak et al. 1998). This led us to
receptor antagonist desegregates eye-specific stripes. Proc Natl Acad Sci
hypothesize that GABA may provide a metabotropic signal
that is important for synapse maturation. In the present study,
HAFIDI A. Distribution of BDNF, NT-3 and NT-4 in the developing auditory
we found that blockade of GABA receptor transduction could
brainstem. Int J Dev Neurosci 17: 285–294, 1999.
eliminate long-lasting synaptic depression (Fig. 1). This result
HAFIDI A, MOORE T, AND SANES DH. Regional distribution of neurotrophin
receptors in the developing auditory brainstem. J Comp Neurol 367: 454 –
is consistent with the ability of a GABA agonist to initiate
long-lasting depression (Fig. 2). While it is not yet clear how
ICHISE T, KANO M, HASHIMOTO K, YANAGIHARA D, NAKAO K, SHIGEMOTO R,
GABA receptor activation initiates inhibitory depression, a G
ATSUKI M, AND AIBA A. mGluR1 in cerebellar Purkinje cells is essential
protein–linked mechanism has recently been shown to depress
for long-term depression, synapse elimination, and motor coordination.
receptor– gated responses through alteration of cy-
Science 288: 1832–1835, 2000.
KANG H AND SCHUMAN EM. Long-lasting neurotrophin-induced enhancement
toskeletal anchoring proteins (Meyer et al. 2000). Postsynaptic
of synaptic transmission in the adult hippocampus. Science 267: 1658 –1662,
GABA receptors apparently exist in LSO since these neurons
exhibited an increased conductance following baclofen expo-
KANG H AND SCHUMAN EM. Intracellular Ca(2ϩ) signaling is required for
sure. However, a presynaptic contribution to inhibitory depres-
neurotrophin-induced potentiation in the adult rat hippocampus. NeurosciLett 282: 141–144, 2000.
sion cannot be ruled out. For example, presynaptic GABAB
KIM HG, WANG T, OLAFSSON P, AND LU B. Neurotrophin 3 potentiates
receptor– coupled mechanisms are known to decrease trans-
neuronal activity and inhibits g-aminobutyratergic synaptic transmission in
mission at both excitatory and inhibitory synapses (Brenowitz
cortical neurons. Proc Natl Acad Sci USA 91: 12341–12345, 1994.
et al. 1998; Lim et al. 2000; Takahashi et al. 1998). However,
KLEINSCHMIDT A, BEAR MF, AND SINGER W. Blockade of “NMDA” receptors
these effects commonly last for seconds to minutes and are not
disrupts experience-dependent plasticity of kitten striate cortex. Science 238:355–358, 1987.
as likely to underlie the long-lasting change we observe in the
KNUDSEN EI AND BRAINARD MS. Visual instruction of the neural map of
auditory space in the developing optic tectum. Science 253: 85– 87, 1991.
Neurotrophins and their receptors have also been implicated
KOMATSU Y. Age-dependent long-term potentiation of inhibitory synaptic
in synapse development and plasticity. In cerebellar cultures,
transmission in rat visual cortex. J Neurosci 14: 6488 – 6499, 1994.
KOTAK VC, KORADA S, SCHWARTZ IR, AND SANES DH. A developmental shift
activity blockade reduces the number of inhibitory synapses,
from GABAergic to glycinergic transmission in the central auditory system.
but inhibitory synaptogenesis is restored by BDNF or neuro-
J Neurosci 18: 4646 – 4655, 1998.
trophin-4 (NT-4), while antibodies to BDNF and NT-4 reduce
KOTAK VC AND SANES DH. Long-lasting inhibitory synaptic depression is age-
inhibitory synapse formation (Seil and Drake-Baumann 2000).
and calcium-dependent. J Neurosci 20: 5820 –5826, 2000.
LEVINE ES, CROZIER RA, BLACK IB, AND PLUMMER MR. Brain-derived neu-
In addition, NT-3 depresses GABA receptor–mediated trans-
rotrophic factor rapidly enhances synaptic transmission in hippocampal
mission in developing cortical neurons (Kim et al. 1994). In the
neurons via postsynaptic tyrosine kinase receptors. Proc Natl Acad Sci USA
MNTB-LSO pathway, immunoreactivity for BDNF, NT-3, and
their receptors is quite prominent during the first two postnatal
IM R, ALVAREZ FJ, AND WALMSLEY B. GABA mediates presynaptic inhibition
at glycinergic synapses in a rat auditory brainstem nucleus. J Physiol (Lond)
weeks (Hafidi 1999; Hafidi et al. 1996). In the present study,
neurotrophin-3 exposure depressed inhibitory synaptic gain
MEYER DM, OLENIK C, HORMANN F, BARTH H, LEEMHUIS J, BR ¨UNIG I,
(Fig. 3A). IPSP amplitude declined within 10 min of exposure,
AKTORIES K, AND N ¨ORENBERG W. Regulation of somatodendritic GABAAreceptor channels in rat hippocampal neurons: evidence for a role of the
but this slow time course may have been due to access to the
small GTPase Rac1. J Neurosci 20: 6743– 6751, 2000.
recording site within the brain slice. The blockade of use-
MOGDANS J AND KNUDSEN EI. Early monaural occlusion alters the neural map
dependent depression by K-252a implies that neurotrophin
of interaural level differences in the inferior colliculus of the barn owl. Brain
receptors may participate along with GABA
MORISHITA W AND SASTRY BR. Chelation of postsynaptic Ca2ϩ facilitates
induce inhibitory depression. One possibility is that the neu-
long-term potentiation of hippocampal IPSPs. Neuroreport 2: 533–536,
rotrophin signal acts to raise intracellular free calcium (Kang
and Schuman 2000), which is required for inhibitory depres-
¨ STBERG AJC, AND VRBOV´A G. Observations on the elimina-
tion of polyneuronal innervation in developing mammalian skeletal muscle.
sion to occur in LSO neurons (Kotak and Sanes 2000). As in
J Physiol (Lond) 282: 571–582, 1978.
excitatory synaptogenesis, adjustments of inhibitory synaptic
ODA Y, KAWASAKI K, MORITA M, KORN H, AND MATSUI H. Inhibitory
strength may thus be regulated by several receptors and intra-
long-term potentiation underlies auditory conditioning of goldfish escape
cellular signaling pathways. Dissection of those mechanisms
behavior. Nature 394: 182–185, 1998.
SANES DH, HARRIS WA, AND REH TA. Development of the Nervous System.
will be critical to appreciate the functionality of inhibitory
synapses before and after sound-evoked activity (Kotak and
SANES DH, KOTAK VC, AND FITZGERALD KK. The developmental influence of
inhibitory synaptic transmission. In: The Biology of Early Influences, editedby Hyson R and Johnson F. New York: Plenum, 2000b.
SANES DH AND SIVERLS V. The development and specificity of inhibitory
This work was supported by National Institute on Deafness and Other
axonal arborizations in the lateral superior olive. J Neurobiol 22: 837– 854,
Communication Disorders Grant DC-00540 to D. H. Sanes.
SANES DH AND TAK ´ACS C. Activity-dependent refinement of inhibitory con-
nections. Eur J Neurosci 5: 570 –574, 1993.
SANES DH AND WALSH EJ. Development of auditory processing. In: Develop-ment of the Auditory System, edited by Rubel EW, Popper AN, and Fay RR.
BRENOWITZ S, DAVID J, AND TRUSSELL L. Enhancement of synaptic efficacy by
presynaptic GABA(B) receptors. Neuron 20: 135–141, 1998.
SCHERER WJ AND UDIN SB. N-methyl-D-aspartate antagonists prevent interac-
CAILLARD O, BEN-ARI Y, AND GAI¨ARSA J-L. Mechanisms of induction and
tion of binocular maps in Xenopus tectum. J Neurosci 9: 3837–3843, 1989.
V. C. KOTAK, C. DIMATTINA, AND D. H. SANES
SEIL FJ AND DRAKE-BAUMANN R. TrkB receptor ligands promote activity-
tion of presynaptic calcium currents and transmitter release by a GABAB
dependent inhibitory synaptogenesis. J Neurosci 20: 5367–5373, 2000.
receptor. J Neurosci 18: 3138 –3146, 1998.
SIMON DK, PRUSKY GT, O’LEARY DDM, AND CONSTANTINE-PATON M. N-
THOMPSON W, KUFFLER DP, AND JANSEN JKS. The effect of prolonged,
methyl-D-aspartate receptor antagonists disrupt the formation of a mamma-
reversible block of nerve impulses on the elimination of polyneuronal
lian neural map. Proc Natl Acad Sci USA 89: 10593–10597, 1992.
innervation of new-born rat skeletal muscle fibers. Neuroscience 4: 271–
TAKAHASHI T, KAHIKAWA Y, AND TSUJIMOTO T. G-protein– coupled modula-
Osteoporosis Screening or Therapy for Osteoporosis for Women Aged 65 Years and Older This measure is to be reported for all female patients aged 65 years and older — a minimum of once per reporting period. Measure description What if this process or outcome of care is not Percentage of female patients aged 65 years and older who have appropriate for your patient? a centra
LifeBridge Church Amplify Student Ministries Health and Permission Form The following must be completed in full before your student can go on any student ministry trip. !Student’s Name __________________________________Grade______ School _________________________ !Address ______________________________________ City_______________ State______ Zip_____________ !Parents’ Names ___________
 RAPID COMMUNICATION
RAPID COMMUNICATION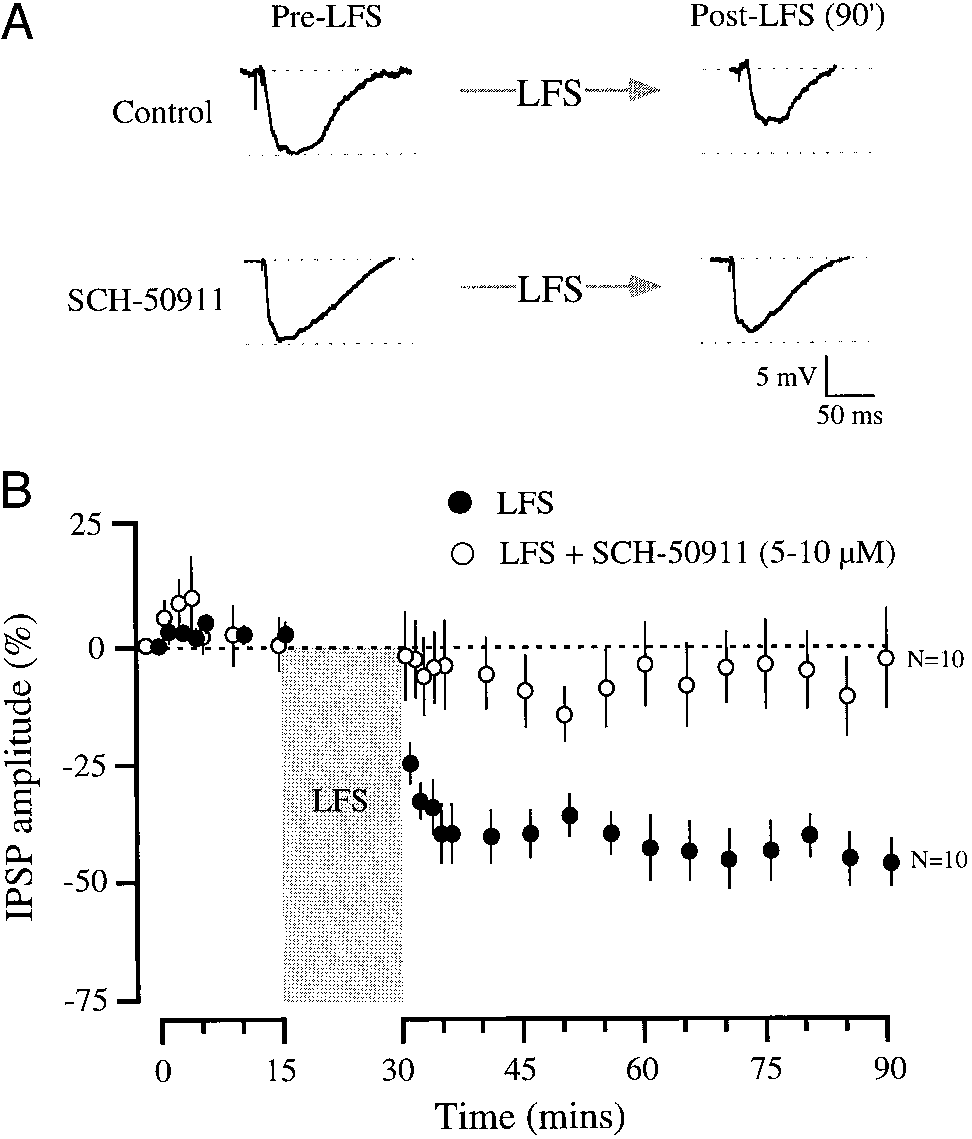 RECEPTOR–MEDIATED INHIBITORY DEPRESSION
neuron, K-252a (200 nM) was added to the internal pipette solution.
RECEPTOR–MEDIATED INHIBITORY DEPRESSION
neuron, K-252a (200 nM) was added to the internal pipette solution.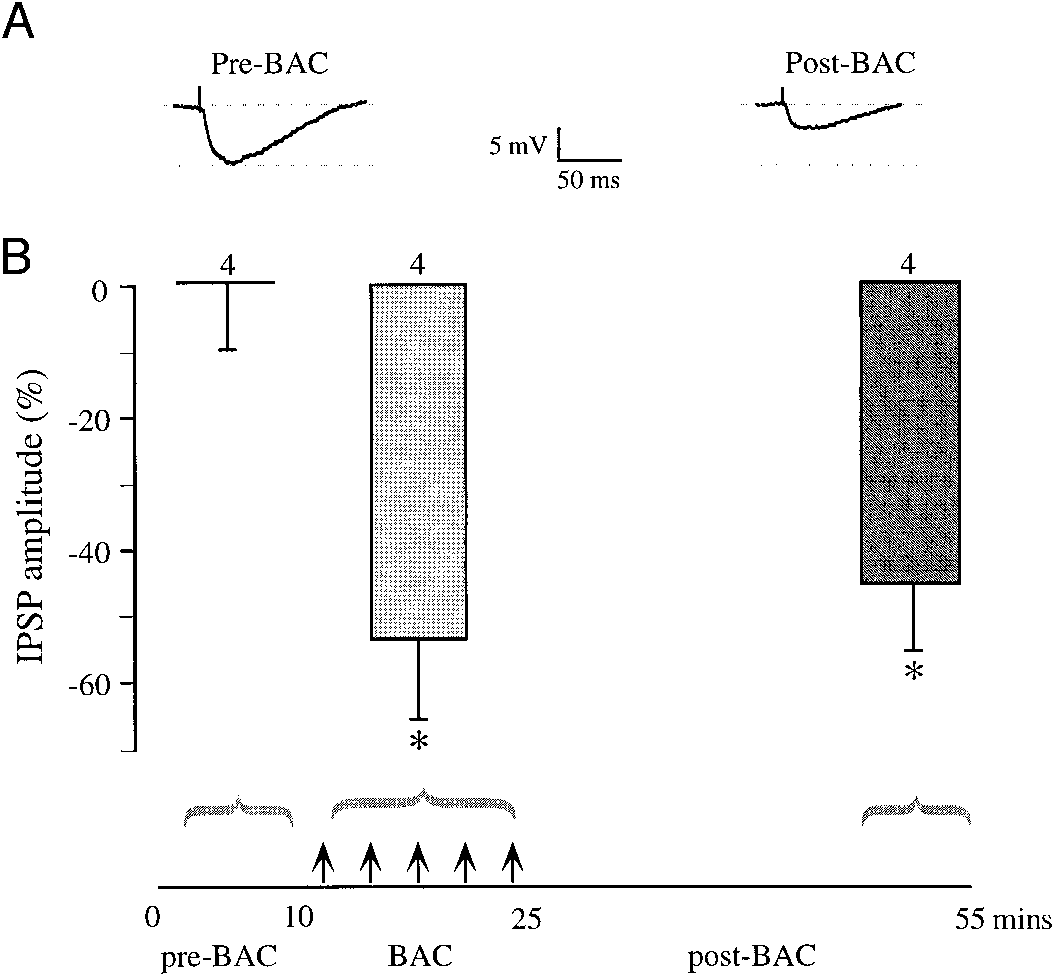
 V. C. KOTAK, C. DIMATTINA, AND D. H. SANES
by both auditory and visual experience (Knudsen and Brainard1991; Mogdans and Knudsen 1993). In the gerbil LSO, inter-aural level difference coding improves with age, and severalanatomical and physiological properties are disrupted by deaf-ferentation during development (Sanes et al. 2000b). We havepreviously shown that inhibitory projections from MNTB toLSO become refined during development, and this process isdisrupted by deafferentation (Sanes and Siverls 1991; Sanesand Taka´cs 1993). More recently, we have found that thestrength of these inhibitory synapses depends on activity, andthis phenomenon wanes with age (Kotak and Sanes 2000). Thepresent results suggest that use-dependent depression of inhib-itory synapses requires GABA receptors, and may also em-
Repeated activation of GABA receptors elicited long-lasting syn-
aptic depression. A: a maximum amplitude IPSP is shown for a P10 neuronbefore and after repeated baclofen exposure. B: the bar graph compares thepercent change in the normalized IPSP amplitude during a control period, a15-min drug exposure period, and after a recovery period. There was asignificant decline (asterisk) in IPSP amplitudes during baclofen treatment, andthis depression persisted at 30 min after the last baclofen exposure. The changein IPSPs was calculated by comparing the average normalized IPSP amplituderecorded during baclofen exposure and at 50 – 60 min of the experiments withthe initial IPSP amplitude (0%) during 1st 10 min of the recording session(comparison between initial IPSPs and IPSPs during baclofen exposure: t ϭ4.01, df ϭ 6, P Ͻ 0.007; comparison between initial IPSPs and IPSPs at 50 – 60min: t ϭ 4.08, df ϭ 6, P Ͻ 0.006).
V. C. KOTAK, C. DIMATTINA, AND D. H. SANES
by both auditory and visual experience (Knudsen and Brainard1991; Mogdans and Knudsen 1993). In the gerbil LSO, inter-aural level difference coding improves with age, and severalanatomical and physiological properties are disrupted by deaf-ferentation during development (Sanes et al. 2000b). We havepreviously shown that inhibitory projections from MNTB toLSO become refined during development, and this process isdisrupted by deafferentation (Sanes and Siverls 1991; Sanesand Taka´cs 1993). More recently, we have found that thestrength of these inhibitory synapses depends on activity, andthis phenomenon wanes with age (Kotak and Sanes 2000). Thepresent results suggest that use-dependent depression of inhib-itory synapses requires GABA receptors, and may also em-
Repeated activation of GABA receptors elicited long-lasting syn-
aptic depression. A: a maximum amplitude IPSP is shown for a P10 neuronbefore and after repeated baclofen exposure. B: the bar graph compares thepercent change in the normalized IPSP amplitude during a control period, a15-min drug exposure period, and after a recovery period. There was asignificant decline (asterisk) in IPSP amplitudes during baclofen treatment, andthis depression persisted at 30 min after the last baclofen exposure. The changein IPSPs was calculated by comparing the average normalized IPSP amplituderecorded during baclofen exposure and at 50 – 60 min of the experiments withthe initial IPSP amplitude (0%) during 1st 10 min of the recording session(comparison between initial IPSPs and IPSPs during baclofen exposure: t ϭ4.01, df ϭ 6, P Ͻ 0.007; comparison between initial IPSPs and IPSPs at 50 – 60min: t ϭ 4.08, df ϭ 6, P Ͻ 0.006).