Research article Nitric oxide is involved in growth regulation and re-orientation of pollen tubes Ana Margarida Prado1, D. Marshall Porterfield2 and José A. Feijó1,3,*
1Instituto Gulbenkian de Ciência, PT-2780-156 Oeiras, Portugal2University of Missouri-Rolla, Department of Biological Sciences, 105 Schrenk Hall, 1870 Miner Circle, Rolla, MO 65409, USA3Centro de Biotecnologia Vegetal, Faculdade de Ciências, Universidad de Lisboa, Campo Grande, Ed.C2. PT-1749-016 Lisboa,Portugal*Author for correspondence (e-mail: jose.feijo@fc.ul.pt)
Development 131, 2707-2714Published by The Company of Biologists 2004doi:10.1242/dev.01153
Summary Nitric oxide (NO) controls diverse functions in many cells pathway. The sensitivity threshold of the response was and organs of animals. It is also produced in plants and significantly augmented by sildenafil citrate (SC), an has a variety of effects, but little is known about their inhibitor of cGMP-specific phosphodiesterases in animals. underlying mechanisms. In the present study, we have NO distribution inside pollen tubes was investigated using discovered a role for NO in the regulation of pollen tube DAF2-DA and was shown to occur mostly in peroxisomes. growth, a fast tip-growing cellular system. Pollen tubes Peroxisomes are normally excluded from the tip of pollen must be precisely oriented inside the anatomically complex tubes and little if any NO is found in the cytosol of that female ovary in order to deliver sperm. We hypothesized region. Our data indicate that the rate and orientation of that NO could play a role in this guidance and tested this pollen tube growth is regulated by NO levels at the pollen hypothesis by challenging the growth of pollen tubes with tube tip and suggest that this NO function is mediated by an external NO point source. When a critical concentration was sensed, the growth rate was reduced and the growth axis underwent a subsequent sharp reorientation, after which normal growth was attained. This response was abrogated in the presence of the NO scavenger CPTIO and affected by drugs interfering in the cGMP signaling
Key words: Pollen, NO, cGMP, Peroxisome, Guidance, ArabidopsisIntroduction
mutant of Arabidopsis, the ovule entrance (mycropile) can
Unraveling the molecular mechanism of pollen tube guidance
direct pollen tube growth over distances of 50-90 μm (Shimizu
is a central issue in sexual plant reproduction and there is a
and Okada, 2000). Recently, the GABA molecule, a
general agreement that directional growth should depend on
neurotransmitter in animals, was proposed to be a part of this
physical and chemical signals that are exchanged between the
navigation system, presumably through the formation of a
male and female gametophytes (Pruitt, 1999; Palanivelu and
continuous gradient towards the ovule that would be sensed
Preuss, 2000; Cheung and Wu, 2001; Johnson and Preuss,
and acted upon by the growing pollen tube (Palanivelu et al.,
2002; Lord and Russel, 2002). Despite intense research efforts
2003). The fact that pollen tube guidance frequently fails in
in the past two decades aimed at defining a mechanistic
crosses between relatively closely related species implies that
explanation of the process, consensus around a central or
at least some of the signals must be species specific. However,
conserved theory of pollen tube guidance is still lacking.
this does not rule out a role for more universal simple
Chemotropic roles have been suggested for diverse style
molecules such as GABA; it may simply mean that the
extracts, sugars, Ca2+, available water, and, more recently,
specificity of the guidance cues comes from differential
short-range effects have been described for lipids,
response or differential sensitivity to a common signal
arabinogalactan-proteins and adhesins (Wolters-Art et al.,
1998; Wu et al., 1995; Mollet et al., 2000) (reviewed by
However, given the biological relevance of fertilization, it is
Johnson and Preuss, 2002). Genetic evidence has also
plausible that evolution has created functional redundancy or
accumulated for long-distance guidance cues, mostly on the
co-functionality for different molecules. In fact, theoretical
basis of mutant screening for reproductive defects (Hülskamp
arguments have been raised that a single chemical gradient
et al., 1997; Ray et al., 1997). In one of the best-characterized
could hardly be responsible for guidance in most species,
physiological models (Torenia fournieri), diffusible signals
which led Lush et al. (Lush et al., 1998) to propose
from the synergid cells of the embryo sac are effective at
mechanical/structural stringencies as co-operative mechanisms
distances of 100-200 μm (Higashiyama et al., 2003). On a
in the guidance of pollen tubes. Classical experiments show
different experimental basis, it was estimated that in the maa
that directionality of growth along the pistil/ovary can in
principle occur in more than one direction, restricting the
NO imaging
guidance cue necessity to just a few crucial steps along the
The presence of NO in pollen tubes was assayed and visualized with
pollen tube path and overruling positive single molecule
10 μM 4,5-diaminofluorescein diacetate (DAF-2DA, Molecular
chemotropism as the sole mechanism of guidance (reviewed by
Probes). Pollen tubes longer than 200 μm were grown in a glass
Heslop-Harrison, 1987; Mascarenhas, 1993; Lord and Russel,
coverslip coated with 0.01% PLL (poly-l-lysine hydrobromide, Mr
331; Sigma), incubated for 5 minutes and perfused to wash excess
In a effort to bridge the gap between in vitro and in vivo
fluorophore. Imaging was carried out using confocal (488 nm) or two-photon excitation (890 nm) on a BioRad MRC1024MP with a
experiments of pollen tube growth manipulation, our attention
Coherent Mira/Verdi Ti-Sa laser, using a Nikon PlanFluo NA1.3 lens.
was drawn to NO as a possible communication molecule in this
Emission was collected with a 522DF35 filter. Images were processed
system on the basis of a number of well-known characteristics
with Metamorph (MM; Universal Imaging Corporation, v. 6.1).
derived from studies in animals (reviewed by Ignarro, 2000;
Kymographs were produced by averaging pixel intensity along a
Stamler, 1994): (1) NO diffuses freely across cell membranes;
linescan of the whole pollen tube at each time point. Pollen tube length
(2) it is known to act as an intra and inter-cellular messenger
is represented on the horizontal axis of the kymograph and time on
in a number of regulation mechanisms; (3) it is known to act
the vertical axis. The tip boundary was aligned on the right side of
as positional cue diffusing from point sources; and (because it
the kymograph by applying a custom-made journal under MM.
is a gas) (4) it acts on minimal thresholds over considerable
NO flux measurements
distances. In plants, NO has been proposed as a regulator of
Carbon fiber microelectrodes were built and operated as previously
growth and developmental processes (Lamattina et al., 2003),
described for NO flux measurements (Cahill and Wightman, 1995;
as exemplified in roots, where NO mediates the response
Friedman et al., 1996; Porterfield et al., 2001). Electrodes were
to indole acetic acid during adventitious root formation
polarized for NO detection at +9.0 V (versus Ag/AgCl half cell
(Pagnussat et al., 2003), in senescence by downregulating
connected to the solution by a 0.5% agarose/3 M KCl bridge) and
ethylene emission (Leshem et al., 1998) and through the
calibrated by dilution of a standard 2 mM NO solution (Gevantman,
stimulation of seed germination (Beligni and Lamattina, 2000).
1995). To characterize the NO gradients created by SNAP, an artificial
NO also promotes adaptive responses against drought stress
NO source (aNOs) was immersed in medium and allowed to reach
operating downstream from ABA (Mata and Lamattina, 2001),
equilibrium. The self-referencing polarographic NO vibrating-
and it has been implicated in the establishment of legume
electrode was stepped linearly from the aNOs tip at 10 μm intervals
Rhyzobium symbiosis (Hérouart et al., 2002). In plant disease
and NO fluxes measured at each point. The diffusion of NO isdescribed by Fick’s Law (J=–DΔC/Δr), where J is expressed as pmol
resistance, NO plays a role by enhancing the induction of
cm–2 s–1, D is the diffusion coefficient for NO (2.6×10–5 cm–2 s–1),
hypersensitive response (Delledonne et al., 1998; Durner et al.,
ΔC is the concentration difference between two electrode positions
and Δr is the excursion path of the electrode (10 μm). The conversion
We present data to indicate that NO can function as a pollen
of the electrode signal to a concentration differential followed a
tube growth modulator by inducing growth re-orientation, the
previously established protocol (Porterfield and Smith, 2000). Data
crucial cellular response to pollen navigation on the pistil.
acquisition, processing and control of electrode movements were
Pollen tubes respond to threshold concentrations of NO by
accomplished using a 3D stepper micropositioner and amplifier
sharp re-orientation, and this reaction is totally abrogated by
(www.applicableelectronics.com) controlled by ASET software
adding the NO scavenger CPTIO to the medium. Furthermore,
(www.ScienceWares.com) (Shipley and Feijó, 1999).
we provide data to indicate that this response is mediated
Subcellular characterization
through a cGMP pathway, and that NO is primarily
Pollen tubes loaded with DAF2-DA were co-incubated with specific
synthesized in peroxisomes. On the basis of these data, we
probes for mitochondria (Rhodamine 1,2,3; 10 μM, Molecular
propose an NO-based regulatory growth mechanism that could
Probes), acidic organelles (LysotrackerRed, 100 μM, Molecular
account for the basic curvature needed for ovule targeting by
Probes) and Golgi (Bodipy-TR, 1 μM, Molecular Probes) and
observed by confocal microscopy. Peroxisomes were imaged bytransient expression of an ECFP-Peroxi construct (6931-1, Clontech). This vector contains a fusion between ECFP and the peroxisomal
Materials and methods
targeting signal 1 (PTS1), which was extracted and cloned into aconstruct containing the LAT52 pollen-specific promoter (Twell et al.,
Pollen germination and tube growth NO assays
1990) in a pBluescript SKII vector (Chen et al., 2002). Tungsten
Fresh Lilium longiflorum pollen was germinated in 1.6 mM
particles (1.1 μm, BioRad, Hercules, CA) were coated with this
H3BO3,1.0 mM KCl, 500 μM CaCl2, 6% sucrose and 50 μM MES,
construct and bombarded into Lilium pollen using the biolistic
pH 6. Healthy, growing tubes were challenged with an artificial NO
PDS-1000/He system (BioRad). After bombardment, pollen was
source (aNOs), a micropipette (20-30 μm tip diameter), tip-filled with
germinated in coverslip-bottom Petri dishes coated with 0.01%
1% agarose and 10 mM SNAP (s-nitroso-acethilpenilcilamine,
poly-l-lysisine hydrobromide, Mr 331 and imaged 10 hour after
Sigma). Control for reactive oxygen intermediates was done carried
germination with a Leica Confocal microscope TC-SP2/AOBS
out by addition of 100 U ml–1 SOD+Catalase. NO specificity was
(excitation at 458 nm to a spectral gate ranging from 469 to 500 nm).
assayed by perfusion of 200 μM of the NO-scavenger CPTIO [2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide, K+-salt, Calbiochem] in presence of an aNOs. Sildenafil citrate (SC;
ViagraTM, Pfizer) was assayed from 319 to 339 μml–1 with a diluted
Lily pollen tubes show a negative tropic response in
aNOs (5-5.8 mM SNAP in 1% agarose). Randomly chosen pollen
the presence of a nitric oxide point source
tubes were challenged with diluted aNOs. Absence of re-orientationresponse was interpreted as indicating a sub-threshold sensitivity of
Pollen tube growth is restricted to the pollen tube tip (Feijó et
that pollen tube to the imposed gradient. Under these conditions, SC
al., 2001). To address if NO plays any effect on Lilium
was perfused and the response recorded by time-lapse video. longiflorum pollen tube growth we designed an in vitro system
NO-mediated regulation of pollen tube growth 2709
to deliver NO specifically to the tip. A glassmicropipette was loaded with an agarose solutioncontaining the NO donor SNAP (s-nitroso-acetylpenicillamine), allowing the molecule to diffuseto the medium and establishing a gradient in thevicinity of the growing pollen tube tip. The pointdiffusion gradient was allowed to settle in liquidgermination medium and the growing pollen tubeswere then placed 60 μm away facing the pipette tip andgrowth was recorded by time-lapse videography. Aspollen tubes move into the gradient, their growth isreduced or, in some cases, completely abrogated. After12-15 minutes pollen tube growth resumes, but with thegrowth axis sharply rotated by an average angle of97.7±3.6° (mean±s.e.m.; n=28), an angle that is similarto the curvatures observed when pollen tubes targetovules in lily (Janson et al., 1994) and Arabidopsis(Shimizu and Okada, 2000), but remarkably sharperthan that produced by any other treatment to lilly pollentubes. After the new growth axis is re-established,pollen tubes achieve a normal growth rate (Fig. 1A). The same pollen tube could be induced to re-orient itsgrowth axis several times by repeated exposure toexogenous NO (Fig. 1B; arrows indicate the position ofthe source). The formation of an NO gradient by SNAPwas confirmed by measurement with a self-referencingvibrating polarographic microelectrode selective forNO. A typical exponential-decay diffusion field from
Fig. 1. (A) Time-lapse sequence of a Lilium longiflorum (lily) pollen tube
the point source was observed (Fig. 1C). These
growing facing an extracellular NO point-source (SNAP on agarose; left on
measurements indicate that the extracellular activation
the image). Pollen tube slows as it moves into the NO-gradient, but direction
threshold for the reorientation response is on the order
proceeds unchanged for ~12 minutes. A new growth axis then starts to be
of 5-10 nmol l–1, or a flux of 0.1-0.2 pmol cm–2 s–1,
defined, forming a sharp right angle from the original axis (97.7±3.6°, n=28).
values well within the physiological range of NO action
The pollen tube then regains normal growth rate (16-20 minutes). Scale bar:
(Ignarro, 2000; Lamattina et al., 2003). The re-
30 μm. (See Movie 1 at http://dev.biologists.org/supplemental) (B) Lily
orientation was maintained after addition of catalase
pollen tube showing three consecutive re-orientation responses induced bymoving the same source to the locations marked with arrows. The growth
and superoxide dismutase (SOD), excluding the
axis changed reproducibly by right angles after each challenge by the NO
possibility of chemical reactions between NO and
source in front of the pollen tube tip. (C) Artificial NO source measurements
reactive oxygen species (ROS), and the subsequent
using a vibrating self-referenced polarographic probe to NO. The graph
secondary production of NO-derived molecules, such
shows a typical exponential NO gradient decay from the point source at
different step distances. Although variations between sources were detected,
More importantly, the re-orientation was totally
these measurements show that within the effective distance (see A) the NO
abrogated by addition of the specific NO scavenger
concentration is in the range of 5-10 nmol l–1, and the NO flux is in the range
CPTIO, a condition in which pollen tubes were
of 0.1-0.2 pmol cm–2 s–1 (values well within the physiological range accepted
observed to grow at normal rates inside the SNAP-
for NO action). (D) Time-lapse sequence of a pollen tube being challenged
containing pipette (intracellular data on the CPTIO
with a diluted NO artificial source in the presence of sildenafil citrate(ViagraTM) (numbers in the top right-hand corner represent minutes after
detection of the response). Using these diluted sources, most pollen tubes do
cGMP mediates pollen tube re-orientation
not show any response, often growing into the pipette. For this experiment,pollen tubes were first incubated on standard medium and challenged with the
response
diluted NO source. If a pollen tube showed no response, i.e. if it was
The re-orientation of the growth axis in the presence of
demonstrated to be insensitive to such low amounts of NO, the medium was
an external gradient lead us to further investigate the
perfused with sildenafil citrate and the same pollen tube is challenged with
existence of downstream messengers of NO. In animal
the same NO source. Despite the lower amount of NO, reverse re-orientation
cells, NO effects can be mediated by cGMP-
angles were observed in the presence of sildenafil citrate (109.8± 9.8°, n=9)
independent signaling pathways (Ignarro, 2000), and it
showing a sensitization effect, from unresponsive to the peak response (see
is well established that this second messenger conveys
movie 1 at http://dev.biologists.org/supplemental).
NO signaling in a number of physiological conditions. cGMP levels are modulated by NO in animal cells, and
number of described effectors of its activity: IBMX, a general
equilibrium concentrations of cGMP are dependent on NO-
inhibitor of the PDE family, and sildenafil citrate (SC;
activated guanylate cyclases (GC) and breakdown activity of
VIAGRATM), a drug that inhibits cGMP degrading
phospodiesterases (PDEs). Although these enzymes have not
phosphodiesterases (PDE5 and PDE6) in mammals (Corbin
been well characterized in plants, we tested the effects of a
and Francis, 1999). The use of IBMX at different
concentrations promoted the occurrence of diverse tip
exogenous NO on pollen tube growth is mediated by cGMP,
abnormal or subnormal morphologies, pointing to the
and that this second messenger is in the signaling cascade that
occurrence of pleiotropic effects (data not shown). Although
affects the growth regulation mechanism.
the drug clearly disrupted growth regulation, the pleiotropicresponses made it difficult to isolate or test its specificity to the
Nitric oxide is produced in peroxisomes
Once a re-orientation response takes place, we asked whether
SC, however, has recently been shown to delay flower
this means that the extracellular NO challenge induces this
senescence (Leshem, 2000). If the re-orientation response of
response by modulating the intracellular NO levels. To address
pollen tubes to exogenous NO involves cGMP and if SC
this question, we searched to see if NO was endogenously
inhibited cGMP degradation in plants, the drug should
produced in pollen tube. Live cells were loaded with the
sensitize pollen tubes to NO and/or prolong the re-orientation
NO sensitive fluorophore, DAF2-DA (4,5-diaminofluorescein
response. To test this possibility we exposed pollen tubes to
diacetate) and imaged by confocal or two-photon microscopy
suboptimal doses of SNAP. The criterion to validate this
(Fig. 2). This probe was previously shown to be NO-specific
experiment was the detection of pollen tubes insensitive to a
in plant tissues (Foissner et al., 2000). Fluorescence was found
less concentrated NO source. Thus, pollen tubes that were
throughout the cytosol, although in the region subjacent to the
previously shown not to re-orient its growth axis on a
tip it was very low (Fig. 2, 1′). A very strong signal was found
lower SNAP concentration were submitted to a SC final
in round organelles of about 2 μm diameter (Fig. 2, 1′). The
concentration of 339 μM. Under these conditions, eight out of
spatiotemporal dynamics of intracellular NO (iNO) are shown
nine pollen tubes (n=9) showed re-orientation after addition of
in the form of kymographs in which we averaged an active
SC, and some even re-oriented to previously never observed
representative region inside each pollen tube at each time-point
angles of 180° (Fig. 1D). We further confirmed this result by
as a color-coded line, and plotted these lines as a function of
measuring an increase of the average re-orientation angle to
time (YY′ axis) and pollen tube length (XX′ axis) (Fig. 2). For
120°±12 (n=9). This angle is 25% steeper than the control but,
the sake of clarity, the pollen tube tips were right-side aligned,
more significantly, is obtained with much lower concentrations
and therefore the slope on the left side of the kymograph
of NO. Dose-response curves of SC also showed a dose-
reflects the pollen tube growth rate. In non-challenged pollen
dependent stimulatory effect of ~30% on the growth rate of
tubes, no significant variation over time is seen. NO levels are
pollen tube at 50 μM. This finding suggests that the effect of
very low in the tip and highest in the subapical domain(control; Fig. 2). To validate the specificity of the observedsignal, the NO scavenger CPTIO [2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide] was applied in
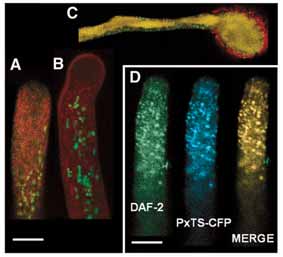
Fig. 2. Detection of intracellular NO in a growing pollen tube of lily using the NO-specific fluophore DAF2-DA (1′). Fluorescence is seen in the cytosol, with less intensity in the apical domain, and is very bright on round cytoplasmic organelles. The spatiotemporal dynamics of intracellular NO is shown in the form of kymographs in which we averaged an active representative region inside each pollen tube at each time-point as a color-coded line (see top wedge), and plotted these lines as a function of time (y-axis) and pollen tube Fig. 3. The DAF-2DA-positive organelles are peroxisomes.
length (x-axis). For the sake of clarity, the pollen tube tip was aligned
(A-D) Confocal images of growing lily pollen tubes incubated in
with the right side, and therefore the slope on the left side of the
DAF-2DA (green) and in organelle-specific dyes (red). No co-
kymograph reflects the growth rate. The chronological order of each
localization was found in the endomembrane system (A, BodipyTR),
time point is read from top to bottom as illustrated by the arrow on
mitochondria (B, Rhodamine 123) or acidic organelles (C,
the y-axis. In a non-challenged pollen tube (control), no significant
LysotrackerRed). (D) Peroxisomes were then tagged by transient
variation along time is seen. Apical depletion and subapical
transformation of pollen grains with a construct containing the
accumulation of NO are clearly visible. Incubation with the NO-
pollen-specific LAT52 promoter driving an ECFP-peroxisome
scavenger CPTIO (44′) almost suppressed the signal from the
targeting signal (PxTS) fusion. Pollen tubes were observed 10 hours
cytosol, but the round organelles are still distinguishable. Kymograph
after germination. The NO-producing organelles (DAF-2, green)
analysis shows the overall decrease after CPTIO addition, but the
show an almost complete co-localization with the ECP signal (PxTS-
apical/subapical pattern, polarity and dimensions are maintained.
CFP, blue), as shown in the merged image. Scale bars: 15 μm.
NO-mediated regulation of pollen tube growth 2711
continuous perfusion. The fluorescencesignal was always reduced in tubes thatwere perfused with CPTIO. Typically,cytosolic NO was almost completelyeliminated, confirming the presence of NOinside pollen tubes and the specificity ofthe probe. Even though the cytosolic NOsignal decayed, the round organelles stillshowed a clear signal even after 44minutes, suggesting that they continued togenerate NO at a high rate (Fig. 2, 44′and lower kymograph). These brightlyfluorescent organelles visible after DAF2-DA exposure have a diameter of about 2
μm (2.17±0.17). We performed double-labeling experiments using differentorganelle-specific probes and DAF-2DAprobe to determine their identity. Doublelabeling with DAF2-DA (green) andselective, color complementing dyes (red)for Golgi and ER (Bodipy-TR, Fig. 3A),mitochondria (rhodamine 123, Fig. 3B)and acidic organelles (Lysotracker, Fig. 3C) showed no co-localization. Asperoxisomes and plastids are still withinthis size range, we transiently transformedpollen tubes with a peroxisomal targetingsignal (PxTS) fused to ECFP under thecontrol of the pollen-specific promoterLAT52. LAT52 is a very late-actingpromoter in monocots, and thus we imagedpollen tube 10 hours after germination. Thedouble labeling with ECFP and DAF2-DAco-localized to the same organelles (Fig. 3D) and morphometric analysis (diameter
Fig. 4. An increase in intracellular NO precedes re-orientation. The time-lapse sequence
and circularity index) indicated that the
(A-C) shows the changes in intracellular NO after challenge with an extracellular point
source, as reported by DAF-2DA fluorescence. In A, a DAF-2DA loaded pollen tube was
followed for 4 minutes. The inserted kymograph shows the typical NO pattern, and no
significant variation with time. (B) Challenging with an external source produces a rise in
fluorescence within 1 minute. After ~10 minutes, the low NO concentration domain
disappears (arrow) simultaneously with growth arrest (slope on the left side of the
the tip (Feijó et al., 2001; Hepler et al.,
kymograph) and soon after the NO concentration peaks. (C) As the concentration
stabilizes, the negative NO tip gradient starts to be defined and re-orientation occurs. Scale
bars: 15 μm. (D) The average pixel intensity variations of the DAF-2DA signal plotted as afunction of time at the tip of a growing pollen tube before (yellow) and after extracellular
NO challenge (white). Accumulation of intracellular NO is obvious soon after the pollen
tube moves into the gradient, but builds up strongly from a threshold point. When the peak
point is reached, growth is arrested. As soon as growth is regained in the new axis, the level
of NO drops to a stable value, which is about twice that seen before challenge. Addition of
the NO-scavenger CPTIO totally inhibited the re-direction response as illustrated in E-G.
A DAF-2DA stained tube (E) was challenged with an extracellular NO point source in the
presence of CPTIO. While the signal decreased as in Fig. 2, the growth of the tube slowed
but slowly regained normal growth without any change of direction (F). Evolution of
intracellular NO shows that after some initial increase, this reaction is immediately
followed by a decrease to levels below the initial level (G and inserted kymograph). Scalebar: 16 μm.
allowed to grow for 4 minutes, in order toadjust the basal signal magnitude (Fig. 4A)and then brought to the vicinity of the NO source (Fig. 4B).
discernable (arrow in the kymograph). Approximately 10
The re-orientation response is preceded by an increase of NO
minutes after challenge with the NO donor, there is an overall
in the tip region and a reduction in the pollen tube growth rate
sustained increase of the signal, coincident with growth arrest
(Fig. 4C and inserted kymograph). During this period, the
(arrow). The re-orientation response then takes place and
subapical region of low NO concentration is no longer
growth resumes at a normal rate, but iNO remains at higher
levels than before the extracellular challenge with the NO
form (NOS1) with no sequence similarities to any mammalian
source (Fig. 4D). Interestingly, growth is preceded by the re-
isoform (Guo et al., 2003). Consistent with our data, the nos1
definition of the subapical NO-depleted domain. Fig. 4D
mutants show a reduced reproductive growth and fertility,
traces the evolution of NO along the whole sequence (yellow
indicating that NO might participate in these events.
points plot values before NO challenge). Diffusion of NO is
We also show that NO is synthesized in the peroxisomes, a
traceable by the fluorescent signal increase, and growth arrest
hypothesis previously proposed in plants (Barroso et al., 1999;
occurs when the maximal cytosolic values are reached. More
importantly, the re-orientation and re-start of normal growth
Our findings indicate that NO is a negative regulator of
are concomitant with a sharp decrease of NO.
growth, at least in pollen tubes. We show that endogenously
A possible explanation for this could lie on the following
generated NO is low or absent at the tip of pollen tube, but is
sequence of events: NO from the point-source diffuses into the
present at higher levels behind the tip, where the presumed NO-
tip, causes growth arrest and stimulates endogenous NO
generating peroxisomes are located. We suggest that this
production in this region. Thus, much of the NO measured
pattern of NO production is permissive for pollen tube growth
would be endogenously produced, and subsequently decreased
at the tip and may, in the absence of an exogenous NO source,
upon resumption of normal growth. Based on the fluorescence
act as a positive feedback reinforcement of elongation in a
intensity, after growth is resumed, the basal level of iNO
straight growth axis. The ubiquitous presence of NO in the
remained stable around a value that is, on average, twice the
medium, by addition of SNAP, prevents germination and
pixel-intensity than before the response. This change suggests
inhibits pollen tube growth rate. However, pointed application
a different steady-state condition at the end of the process,
of an NO donor near the pollen tube tip results in transient
which could underlie some sort of molecular memory to the
growth arrest, which is followed by re-directed growth. This
first exposure to external NO. Whatever the reason, pollen
effect is mediated by NO that diffuses into the tip. Growth
tubes remain sensitive to NO and are capable of re-direction
arrest is then associated with changes in cell polarity and
again and again (Fig. 1B). Therefore, if some memory is
peroxisome redistribution, as shown by the extension of the
retained, it must be mediated by an increase in sensitivity,
streaming lanes into the tip. We assume that it is the
which could be dependent on higher background levels of
localization of peroxisomes and thus the site of endogenous
NO production, which eventually determines the direction into
We also monitored the NO levels after perfusion with
which pollen tube growth resumes. Peroxisomes are the plant
CPTIO (Fig. 4E-G). Challenge with the NO-source provoked
cell oxidative organelles and there are reports that highlight
a slight increase in NO but immediately after, it decreased
the relevance of this organelles in the production of signal
progressively to levels bellow the initial basal concentration.
molecules, i.e. NO and reactive oxygen species. These
This level then remained constant (inserted kymograph). Under
organelles have a rich enzymatic machinery and are
these conditions, no re-orientation took place, and growth rate
reported to participate in developmental processes such as
slowly recovered. Together these results show a direct
photomorphogenesis in Arabidopsis (Hu et al., 2002; Barroso
relationship between iNO and the regulation of cell growth and
et al., 1999; del Rio et al., 2002).
Although our data show that endogenous NO production is
correlated with the regulation of pollen tube growth, the in vivoconfirmation of this is made difficult by various experimental
Discussion
obstacles. Real-time imaging of pollen tube guidance in vivo
Our data prompt us to propose a new role for NO in plant
would imply the possibility of optical sectioning closed
biology. NO has been previously suggested to act as messenger
flowers, which implies demanding technical conditions (two-
in plant developmental processes, stress, defense responses and
photon excitation and water-immersion, long working distance
symbiosis establishment (Lamattina et al., 2003). We directly
objectives) far from optimized for this specific application.
demonstrate a role for NO in pollen tube growth and guidance.
Excitation-derived photo-damage of cells or, more
In animals, NO is synthesized by NO synthases (NOS) and its
dramatically, any sort of ovary dissection or injury of the ovary
signaling function is mediated, at least in part, by soluble
is not an option because it will generate stress-induced bursts
guanylyl cyclases (sGC) that generate cGMP and by
of NO production (Lamattina et al., 2003). A possibility for
phosphodiesterases (PDEs) that hydrolyze cGMP (Ignarro,
overcoming these obstacles will be either the use of pollen
2000). In plants, no sequences homologous to mammalian
tubes expressing highly fluorescent reporter genes to closely
NOS, sGC and PDEs have been found namely in the
monitor the pollen tube-pistil interaction, or, otherwise, the use
Arabidopsis genome. Yet, there is evidence for cGMP function
of floral mutants with open ovaries and exposed, yet functional,
in plants (Penson et al., 1996; Durner et al., 1998). In
agreement with a role for cGMP in plants, one functional plant
Another problem is related to the high reactivity of NO, with
guanylate cyclase was identified in Arabidopsis, though it
an half life depending on the redox status of the surrounding
shows an unusual domain organization (Ludidi and Gehring,
environment, namely when ROS are present (Ignarro, 2000;
2003). There are also reports of NO production (Barroso et al.,
Thomas et al., 2001). This makes it difficult to gauge the
1999; del Rio et al., 2002). The enzyme nitrate reductase
amount of NO being produced in vivo, so no invasive
appears to produce NO necessary for stomata closure
techniques for NO can easily quantify a putative signal from
(Lamattina et al., 2003). More importantly, two recent reports
the female tissue. A self-referencing NO selective electrode
describe new enzymes with NOS activity, one identified as an
could be used, but again tissue accessibility is a limiting factor.
inducible NOS with a sequence variant to a glycine
Several difficulties arise when interpreting chemical cues
decarboxylase (Chandok et al., 2003) and a constitutive NOS
identified in different plant species: it can be argued that
NO-mediated regulation of pollen tube growth 2713
general mechanisms do not assure species specificity to avoid
the gradient is maintained in time as a single pollen tube can
widespread cross-fertilization (Johnson and Preuss, 2002). One
undergo consecutive re-orientation responses with the same
possible explanation could be related to different threshold
NO source. In addition, the response can be prevented if the
sensitivities operating for a given molecule from species to
gradient is perturbed or annihilated by an NO scavenger.
species. Otherwise a different species could use similar
The events downstream of NO seem to be, at least in part,
mechanism, but with derivatised molecules within a single
mediated by cGMP. Support for this assertion comes from our
chemical family, which would be transduced into different
finding that, among other tested chemicals, sildenafil citrate, a
effects. Given the diversity of molecules shown to have
drug that inhibits cGMP-selective PDEs of mammals,
guidance effects on pollen tubes, and predicting that more will
facilitated the redirected growth of pollen tubes in response to
be uncovered through successive genetic screens, it is likely
low doses of NO donors that were themselves ineffective.
that chemical signaling between the pollen tube and pistil could
Previous studies with cyclic nucleotide analogues also suggest
convey specificity by using universal molecules in various
that cGMP and cAMP are involved in pollen tube growth
control (Moutinho et al., 2001; Elias et al., 2001). A likely
This finding indicates the need for further cues from the
target downstream of cGMP is a family of cyclic nucleotide-
pistil. Whether or not NO takes part in communication cannot
gated channels (CNGs) (Leng et al., 1999), also represented in
be deduced from in vitro studies, but the striking re-orientation
the pollen transcriptome (Becker et al., 2003). Directly or
response warrants for further investigation. In the context of
coupled with other transporters, CNGs may regulate the flux
our data, a feasible NO-guidance mechanism would be
of ions such as Ca2+, H+ and Cl that are known to be involved
possible if there were specialized female tissues acting as NO
in pollen tube growth control (Feijó et al., 2001; Becker et al.,
‘hot spots’, for example, at the base of the funiculus, where a
2003; Feijó et al., 1999; Zonia et al., 2002). Cyclic nucleotide
sharp change in pollen tube growth direction is required, or
balance, modulation of Ca2+ channels and the control of nerve
near the embryo sac after fertilization, in order to prevent
growth bi-directional axon guidance have recently been linked
secondary pollen tube from penetrating the micropyle. The
(Nishyama et al., 2003). These findings encourage further
indication that nos1, the only bona fide NO-producing mutant
efforts to characterize the various components of the NO signal
so far described, shows fertility deficiencies is a positive
pathway in plants and to endue in genetic and biochemical
indication that NO may be involved in pollen tube guidance.
In past research, pollen tube guidance could not be fully
explained by the actions of positive guidance cues. In addition,
We thank A. M. Shipley (www.ApplicableElectronics.com,
it remains debatable that tracking down a molecule will
Foresdale, MA) for vibrating probe support; Alice Cheung (University
overcome questions related to pollen tube path length and
of Massachusetts, Amherst) for the LAT52 promotor expressionvector; Ueli Grossniklaus (University of Zürich), Sheila McCormick
thickness (Lush et al., 1998). Yet, a gaseous molecule may
(USDA Plant Gene Expression Genomics Center, Berkeley) and
overcome these barriers easily. In proposing NO, a diffusible
Sukalyan Chaterjee (IGC) for help and discussion; Nuno Moreno
gas, as a candidate for pollen tube guidance, we may address
(IGC) for imaging support; Werner Haas (IGC) and Tony Trewavas
a controversial aspect of pollen tube guidance. In Arabidopsis,
(University of Edinburgh) for careful critical reading of the
Hülskamp et al. (Hülskamp et al., 1995) propose that each
manuscript. A.M.P. acknowledges an FCT PhD Fellowship
ovule guides the pollen tube by chemotatic gradients with ~100
(SFRH/BD/6278/2001) and a FLAD (Luso-American Foundation
μm range of action at the junction of the ovule with the
for Development) travel grant. J.A.F.’s laboratory is supported by
placenta. However, wild-type Arabidopsis pollen tubes make a
FCT grants POCTI/BCI/46453/2002, POCTI/BCI/41725/2001 and
sharp turn to enter the mycropyle in 10 μm of this area
POCTI/BIA/34772/1999. Purified sildenafil citrate (Viagra™) was a
(Shimizu and Okada, 2000). Surface localized diffusion of
chemotatic signals effective through 50 cells diameters wouldrequire signal molecules of less than one kDa (Ray et al., 1997;
References
Crick, 1970). The ability of NO to function as a messengeracross cell layers and to trigger cellular processes is nowadays
Barroso, J. B., Corpas, F. J., Carreras, A., Sandalio, L. M., Valderrama,
well established in animals (Ignarro, 2000). The negative
R., Palma, J. M., Lupiáñez, J. A. and del, Río, L. A. (1999). Localization of nitric oxide in plant peroxisomes. J. Biol. Chem. 274, 36729-36733.
chemotropism described here for NO is reminiscent of the
Becker, J. D., Boavida, L. C., Carneiro, J., Haury, M. and Feijó, J. A.
effects of semaphorins on axon guidance in animals: these
(2003). Transcriptional profiling of Arabidopsis tissues reveals the unique
proteins function as chemorepellents, which prompt axons to
characteristics of the pollen transcriptome. Plant Physiol. 133, 713-725.
make right angle turns within an environment that contains
Beligni, M. V. and Lamattina, L. (2000). Nitric oxide stimulates seed
germination and de-etiolation, and inhibits hypocotyl elongation, three light-
both attractants and repellents (Tessier-Lavigne and Goodman,
inducible responses in plants. Planta 210, 215-221.
1996). Similarly, NO acts as negative effector on the retinal
Cahill, P. S. and Wightman, R. M. (1995). Simultaneous amperometric
patterning of the optical lobe in Drosophila, where NO
measurement of ascorbate and catecholamine secretion from individual
prevents further extension of axons beyond their target neurons
bovine adrenal medullary cells. Anal. Chem. 67, 2599-2605.
(Gibbs and Truman, 1998). NO function as a guidance cue
Chandok, M. R., Ytterberg, A. J., van Wijk, K. J. and Klessig, D. (2003).
The pathogen-inducible nitric oxide synthase (iNOS) in plants is a variant
implies that (1) it is be able to form a concentration gradient,
of the p protein of the glycine decarboxylase complex. Cell 113, 469-482.
(2) it produces a specific response, (3) it remains stable for a
Chen, C. Y., Wong, E. I., Vidali, L., Estavillo, A., Hepler, P. K., Wu, H. M.
given period of time, and (4) it varies in effectiveness with
and Cheung, A. Y. (2002). The regulation of actin organization by actin-
distance to the target (Palanivelu and Preuss, 2000). Our in
depolymerizing factor in elongating pollen tubes. Plant Cell 14, 2175-2190. Cheung, A. Y. and Wu, H. M. (2001). Pollen tube guidance- right on target.
vitro data support these criteria. We were able to detect an
Science 293, 1441-1442.
artificially generated external NO gradient to which the pollen
Corbin, J. D. and Francis, S. H. (1999). cyclic GMP phosphodiesterase-5:
tubes respond in a specific way (re-orientation growth axis),
target of sildenafil. J. Biol. Chem. 274, 13729-13732. Crick, F. (1970). Diffusion in embryogenesis. Nature 225, 420-422. Ludidi, N. and Gehring, C. (2003). Identification of a novel protein with del, Rio, L. A., Corpas, F. J., Sandalio, L. M., Palma, J. M., Gómez, M.
guanylyl cyclase activity in Arabidopsis thaliana. J. Biol. Chem. 278, 6490- and Barroso, J. B. (2002). Reactive oxygen species, antioxidant systems
and nitric oxide in peroxisomes. J. Exp. Bot. 53, 1255-1272. Lush, W. M., Grieser, F. and Wolters-Arts, M. (1998). Directional guidance Delledonne, M., Xia, Y., Dixon, R. and Lamb, C. (1998). Nitric oxide
of Nicotiana alata pollen tubes in vitro and on the stigma. Plant Physiol.
functions as a signal in plant disease resistance. Nature 394, 585-588. 118, 733–741. Durner, J., Wendehenne, D. and Klessig, F. (1998). Defense gene induction Mascarenhas, J. P. (1993). Molecular mechanisms of pollen tube growth and
in tobacco by nitric oxide, cyclic GMP and cyclic ADP-ribose. Proc. Natl.
differentiation. Plant Cell 5, 1303-1314. Acad. Sci. USA 95, 10328-10333. Mata, C. G. and Lamattina, L. (2001). Nitric oxide induces stomatal closure Elias, M., Cvckova, F., Obermeyer, G. and Zarsky, V. (2001).
and enhances the adaptive plant response against drought stress. Plant
Microinjection of guanine nucleotide analogue into lily pollen tubes results
Physiol. 126, 1196-1204.
in isodiametric tip expansion. Plant Biol. 3, 489-493. Mollet, J. C., Park, S. Y., Nothnagel, E. A. and Lord, E. M. (2000). A lily Feijó, J. A., Sainhas, J., Hackett, G. R., Kunkel, J. G. and Hepler, P. K.
stylar pectin is necessary for pollen tube adhesion to an in vitro stylar matrix.
(1999). Growing pollen tubes possess a constitutive alkaline band in the
Plant Cell 12, 1737-1749.
clear zone and a growth dependent acidic tip. J. Cell Biol. 144, 483-496. Moutinho, A., Hussey, P. J., Trewavas, A. T. and Malhó, R. (2001). cAMP Feijó, J. A., Sainhas, J., Holdaway-Clarke, T., Cordeiro, S., Kunkel, J. G.
acts as a second messenger in pollen tube growth and reorientation. Proc.and Hepler, P. K. (2001). Cellular oscillations and the regulation of growth: Natl. Acad. Sci. USA 98, 10481-10486.
the pollen tube paradigm. BioEssays 23, 86-94. Nishiyama, M., Hoshino, A., Tsai, L., Henley, J. R., Goshima, Y., Tessier- Foissner, I., Wendehenne, D., Langebartels, C. and Durner, J. (2000). In Lavigne, M., Poo, M. M. and Hong, K. (2003). Cyclic AMP/ GMP- vivo imaging of an elicitor-induced nitric oxide burst in tobacco. Plant J.
dependent modulation of Ca2+ channels sets the polarity of nerve growth-
23, 817-824.
cone inducing turning. Nature 423, 990-995. Friedman, M. N., Robinson, S. W. and Gerhardt, G. A. (1996). o- Pagnussat, G. C., Lanteri, M. L. and Lamattina, L. (2003). Nitric oxide and
Phenylenediamine-modified carbon fiber electrodes for the detection of
cyclic GMP are messengers in the indole acetic acid adventious rooting
nitric oxide. Anal. Chem. 68, 2621-2628.
process. Plant Physiol. 132, 1241-1248. Gevantman, L. H. (1995). In Handbook of Chemistry and Physics, 76th edn Palanivelu, R. and Preuss, D. (2000). Pollen tube targeting and axon
(ed. D. R. Lide), p. 63. Boca Raton, FL: CRC Press.
guidance: parallels in tip growth mechanisms. Trends Cell Biol. 10, 517- Gibbs, S. M. and Truman, J. M. (1998). Nitric oxide and cyclic GMP regulate
retinal patterning in the optic lobe of Drosophila. Neuron 20, 83-93. Palanivelu, R., Brass, L., Edlund, A. F. and Preuss, D. (2003). Pollen tube Guo, F.-Q., Okamoto, M. and Crawford, N. M. (2003). Identification of a
growth and guidance is regulated by POP2, an Arabidopsis gene that
plant nitric oxide synthase gene involved in hormonal signalling. Science
controls GABA levels. Cell 114, 47-59. 302, 100-103. Penson, S. P., Schuurink, R. C., Fath, A., Gubler, F., Jacobsen, J. V. and Hepler, P. K., Vidali, L. and Cheung, A. Y. (2001). Polarized cell growth in Jones, R. L. (1996). cGMP is required for gibberellic acid-induced gene
higher plants. Annu. Rev. Cell. Dev. Biol. 17, 159-187.
expression in barley aleurone. Plant Cell 8, 2325-2333. Hérouart, D., Baudouin, E., Frendo, P., Harrison, J., Santos, R., Jamet, Porterfield, D. M. and Smith, P. J. S. (2000). Characterization of trans- A., van de Sype, G., Touati, D. and Puppo, A. (2002). Reactive oxygen
cellular oxygen and proton fluxes from Spirogyra grevilleana using self-
species, nitric oxide and gluthatione: a key role in the establishment of the
referencing microelectrodes. Protoplasma 212, 80-88.
legume – Rhyzobium symbiosis? Plant. Physiol. Biochem. 40, 619-624. Porterfield, D. M., Laskin, J. D., Jung, S.-K., Malchow, R. P., Billack, B., Heslop-Harrison, J. (1987). Pollen germination and pollen tube growth. Smith, P. J. S. and Heck, D. (2001). Proteins and lipids define the Internatl. Rev. Cytol. 107, 1-78.
diffusional field of nitric oxide. Am. J. Physiol. Lung Cell. Mol. Physiol. Higashiyama, T., Kuroiwa, H. and Kuroiwa, T. (2003). Pollen-tube 281, L904-L912.
guidance: beacons from the female gametophyte. Curr. Opin. Plant Biol. 6, Pruitt, R. E. (1999). Complex sexual signals for the male gametophyte. Curr. Opin. Cell. Biol. 2, 419-422. Hu, J., Aguirre, M., Peto, C., Alonso, J., Ecker, J. and Chory, J. (2002). A Ray, S., Park, S. S. and Ray, A. (1997). A pollen tube guidance by the female
role for peroxisomes in photomorphogenesis and development of
gametophyte. Development 124, 2489-2498. Arabidopsis. Science 297, 405-410. Shimizu, K. K. and Okada, K. (2000). Attractive and repulsive interactions Huang, X., Kiefer, E., von Rad, U., Ernst, D., Foissner, I. and Durner, J.
between female and male gametophyte in Arabiposis pollen tube guidance.
(2002). Nitric oxide and nitric oxide-dependent gene induction in plants. Development. 127, 4511-4518. Plant Physiol. Biochem. 40, 625-631. Shipley, A. M. and Feijó, J. A. (1999). The use of the vibrating probe Hülskamp, M., Schneitz, K. and Pruitt, R. E. (1995). Genetic evidence for
technique to study steady extracellular currents during pollen germination
a long range activity that directs pollen tube guidance in Arabidopsis. Plant
and tube growth. In Fertilization in Higher Plants: Molecular andCell 7, 57-64. Cytological Aspects (ed. M. Cresti, G. Cai and A. Moscatelli), pp. 235-252. Ignarro, J. (2000). In Nitric Oxide. Biology and Pathobiology (ed. J. L.
Heidelberg, Berlin: Springer-Verlag.
Ignarro), pp. 3-380. San Diego, CA: Academic Press. Stamler, J. S. (1994). Redox signalling: nitrosylation and related target Janson, J., Reinders, M. C., Valkering, A. G. M., Vantuyl, J. M. and
interactions of nitric oxide. Cell 78, 931-936. Keijzer, C. J. (1994). Pistil exudates production and pollen-tube growth in Tessier-Lavigne, M. and Goodman, C. S. (1996). The molecular biology of Lilium longiflorum Thunb. Ann. Bot. 73, 437-446.
axon guidance. Science 274, 1123-1160. Johnson, M. A. and Preuss, D. (2002). Plotting a course: multiple signals Thomas, D. D., Liu, X., Kantrow, S. P. and Lancaster, J. R., Jr (2001). The
guide pollen tubes to their targets. Dev. Cell 2, 273-281.
biological life time of nitric oxide: implications for the perivascular
Lamattina, L., Garcia-Mata, C., Graziano, M. and Pagnussat, G. (2003).
dynamics of NO and O2. Proc. Natl. Acad. Sci. USA 98, 355-360.
Nitric oxide: the versatility of an extensive signal molecule. Annu. Rev.Twell, D., Yamaguchi, J. and McCormick, S. (1990). Pollen-specific gene Plant. Biol. 54, 109-136.
expression in transgenic plants: coordinate regulation of two different
Leng, Q., Mercier, R. W. and Yao, W. and Berkowitz, G. A. (1999). Cloning
tomato gene promoters during microsporogenesis. Development 109, 705-
and first functional characterization of a plant cyclic nucleotide gated cation
channel. Plant Physiol. 121, 753-761. Wolters-Art, M., Lush, W. M. and Mariani, C. (1998). Lipids are required Leshem, Y. Y. (2000). Nitric Oxide in Plants Occurrence, Function and Use.
for directional pollen-tube growth. Nature 392, 818-821.
Dordrech, The Netherlands: Kluwer Academic. Wu, H. M., Wang, H. and Cheung, A. Y. (1995). A pollen tube growth Leshem, Y. Y., Wills, R. B. H. and Ku, V. V. (1998). Evidence for the function
stimulatory glycoprotein is deglycosylated by pollen tubes and displays a
of the free radical gas – nitric oxide (NO.) – as an endogenous maturation
glycosylation gradient in the flower. Cell 82, 395-403.
and senescence regulating factoring higher plants. Plant Physiol. Biochem.Zonia, L., Cordeiro, S., Tupy, J. and Feijó, J. A. (2002). Oscillatory chloride 36, 825-833.
efflux at the pollen tube apical tip has a role in growth and osmoregulation
Lord, E. M. and Russell, S. D. (2002). The mechanism of pollination and
and is linked to inositol polyphosphate signaling pathways. Plant Cell 14,
fertilization in plants. Annu. Rev. Cell. Dev. Biol. 18, 81-105.
Progress on Medication Safety Medication Safety Forum Chaired by the Health & Children, Ms Marita Kinsella, improvement projects designed to achieve safer, more effective use of medicines. Three Primary Drivers for Safety 1. Reliable Medication Management Processes in residential care settings (acute hospitals, mental health facilities, 2. Good Co-ordination of Care
SPED Contábil, fiscal e Social - Empresas precisam estar preparadas paras as mudanças - Por Robson Lopes Bezerra SPED Contábil, fiscal e Social - Altere a realidade fiscal. Contábil e Social da sua empresa. Por Robson Lopes Bezerra Confira as mudanças que já estão valendo e as que entrarão em vigor em janeiro de 2012 - Diante de tantas mudanças é importante estes conh
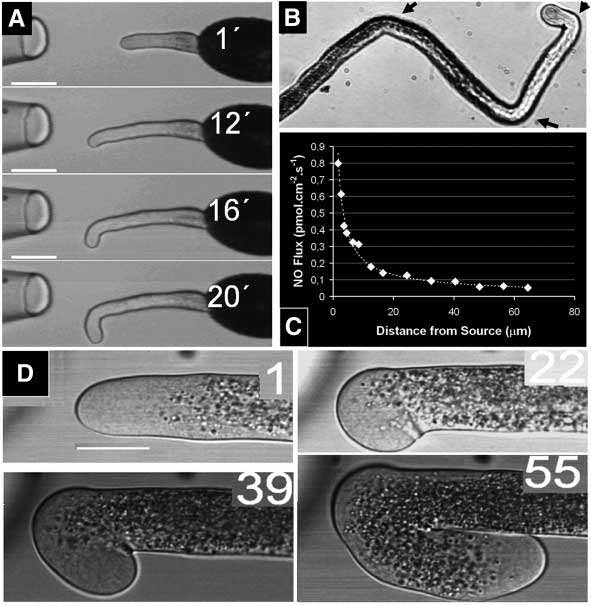 NO-mediated regulation of pollen tube growth 2709
to deliver NO specifically to the tip. A glassmicropipette was loaded with an agarose solutioncontaining the NO donor SNAP (s-nitroso-acetylpenicillamine), allowing the molecule to diffuseto the medium and establishing a gradient in thevicinity of the growing pollen tube tip. The pointdiffusion gradient was allowed to settle in liquidgermination medium and the growing pollen tubeswere then placed 60 μm away facing the pipette tip andgrowth was recorded by time-lapse videography. Aspollen tubes move into the gradient, their growth isreduced or, in some cases, completely abrogated. After12-15 minutes pollen tube growth resumes, but with thegrowth axis sharply rotated by an average angle of97.7±3.6° (mean±s.e.m.; n=28), an angle that is similarto the curvatures observed when pollen tubes targetovules in lily (Janson et al., 1994) and Arabidopsis(Shimizu and Okada, 2000), but remarkably sharperthan that produced by any other treatment to lilly pollentubes. After the new growth axis is re-established,pollen tubes achieve a normal growth rate (Fig. 1A).
NO-mediated regulation of pollen tube growth 2709
to deliver NO specifically to the tip. A glassmicropipette was loaded with an agarose solutioncontaining the NO donor SNAP (s-nitroso-acetylpenicillamine), allowing the molecule to diffuseto the medium and establishing a gradient in thevicinity of the growing pollen tube tip. The pointdiffusion gradient was allowed to settle in liquidgermination medium and the growing pollen tubeswere then placed 60 μm away facing the pipette tip andgrowth was recorded by time-lapse videography. Aspollen tubes move into the gradient, their growth isreduced or, in some cases, completely abrogated. After12-15 minutes pollen tube growth resumes, but with thegrowth axis sharply rotated by an average angle of97.7±3.6° (mean±s.e.m.; n=28), an angle that is similarto the curvatures observed when pollen tubes targetovules in lily (Janson et al., 1994) and Arabidopsis(Shimizu and Okada, 2000), but remarkably sharperthan that produced by any other treatment to lilly pollentubes. After the new growth axis is re-established,pollen tubes achieve a normal growth rate (Fig. 1A).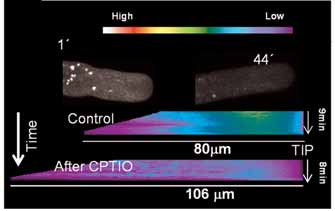
 concentrations promoted the occurrence of diverse tip
exogenous NO on pollen tube growth is mediated by cGMP,
abnormal or subnormal morphologies, pointing to the
and that this second messenger is in the signaling cascade that
occurrence of pleiotropic effects (data not shown). Although
affects the growth regulation mechanism.
concentrations promoted the occurrence of diverse tip
exogenous NO on pollen tube growth is mediated by cGMP,
abnormal or subnormal morphologies, pointing to the
and that this second messenger is in the signaling cascade that
occurrence of pleiotropic effects (data not shown). Although
affects the growth regulation mechanism. NO-mediated regulation of pollen tube growth 2711
continuous perfusion. The fluorescencesignal was always reduced in tubes thatwere perfused with CPTIO. Typically,cytosolic NO was almost completelyeliminated, confirming the presence of NOinside pollen tubes and the specificity ofthe probe. Even though the cytosolic NOsignal decayed, the round organelles stillshowed a clear signal even after 44minutes, suggesting that they continued togenerate NO at a high rate (Fig. 2, 44′and lower kymograph). These brightlyfluorescent organelles visible after DAF2-DA exposure have a diameter of about 2
μm (2.17±0.17). We performed double-labeling experiments using differentorganelle-specific probes and DAF-2DAprobe to determine their identity. Doublelabeling with DAF2-DA (green) andselective, color complementing dyes (red)for Golgi and ER (Bodipy-TR, Fig. 3A),mitochondria (rhodamine 123, Fig. 3B)and acidic organelles (Lysotracker, Fig.
NO-mediated regulation of pollen tube growth 2711
continuous perfusion. The fluorescencesignal was always reduced in tubes thatwere perfused with CPTIO. Typically,cytosolic NO was almost completelyeliminated, confirming the presence of NOinside pollen tubes and the specificity ofthe probe. Even though the cytosolic NOsignal decayed, the round organelles stillshowed a clear signal even after 44minutes, suggesting that they continued togenerate NO at a high rate (Fig. 2, 44′and lower kymograph). These brightlyfluorescent organelles visible after DAF2-DA exposure have a diameter of about 2
μm (2.17±0.17). We performed double-labeling experiments using differentorganelle-specific probes and DAF-2DAprobe to determine their identity. Doublelabeling with DAF2-DA (green) andselective, color complementing dyes (red)for Golgi and ER (Bodipy-TR, Fig. 3A),mitochondria (rhodamine 123, Fig. 3B)and acidic organelles (Lysotracker, Fig.