Multidrug efflux pump overexpression inStaphylococcus aureus after single and multiplein vitro exposures to biocides and dyes
Aure´lie A. Huet,1,3 Jose L. Raygada,2 Kabir Mendiratta,1 Susan M. Seo2and Glenn W. Kaatz1,2
1John D. Dingell Department of Veterans Affairs Medical Center, Detroit, MI 48201, USA
2Department of Medicine, Division of Infectious Diseases, Wayne State University School of
3E´cole Supe´rieure de Microbiologie et Se´curite´ Alimentaire de Brest, Universite´ de Bretagne
Occidentale, Technopoˆle Brest-Iroise, 29280 Plouzane´, France
Biocides and dyes are commonly employed in hospital and laboratory settings. Many of theseagents are substrates for multiple-drug resistance (MDR)-conferring efflux pumps of bothGram-positive and Gram-negative organisms. Several such pumps have been identified inStaphylococcus aureus, and mutants overexpressing the NorA and MepA MDR pumps followingexposure to fluoroquinolones have been identified. The effect of exposure to low concentrations ofbiocides and dyes on the expression of specific pump genes has not been evaluated. Usingquantitative reverse-transcription PCR we found that exposure of clinical isolates to lowconcentrations of a variety of biocides and dyes in a single step, or to gradually increasingconcentrations over several days, resulted in the appearance of mutants overexpressing mepA,mdeA, norA and norC, with mepA overexpression predominating. Overexpression was frequentlyassociated with promoter-region or regulatory protein mutations. Mutants having significantincreases in MICs of common pump substrates but no changes in expression of studied pumpgenes were also observed; in these cases changes in expression of as-yet-unidentified MDRpump genes may have occurred. Strains of S. aureus that exist in relatively protectedenvironments and are repeatedly exposed to sublethal concentrations of biocides can developefflux-related resistance to those agents, and acquisition of such strains poses a threat to patients
treated with antimicrobial agents that are also substrates for those pumps, such as ciprofloxacin
to contribute to hospital-acquired infections, may berepeatedly exposed to sublethal concentrations of biocides,
Biocides are commonly used in the processes of cleaning
increasing the possibility of the development of reduced
hospital, laboratory and home environments, sterilization
susceptibility (Boyce, 2007; Smith et al., 2008). The
of medical equipment, and skin decontamination prior to
presence of biocide residues on the skin or inanimate
surgery (McDonnell & Russell, 1999; Russell, 2003).
objects may also provide such selective pressure.
Pathogenic organisms in the hospital environment maybe sheltered from the effect of these compounds by the
One mechanism by which reduced biocide susceptibility
concomitant presence of materials that interfere with their
can occur is the upregulation of multiple-drug resistance
action or their existence in an environmental niche that
(MDR) efflux pumps that include these compounds within
provides some protection (McBain et al., 2002; McDonnell
their substrate profiles. While it is agreed that increases in
& Russell, 1999). Such organisms, which have been shown
biocide MICs conferred by efflux pumps probably do notconfer true resistance at the concentrations deployed for
Abbreviations: AF, acriflavine; BAC, benzalkonium chloride; CET,
environmental or cutaneous disinfection, upregulation of
cetrimide; CHX, chlorhexidine; CV, crystal violet; DEQ, dequalinium;
MDR pumps as a result of biocide exposure may provide a
EB, ethidium bromide; MDR, multiple-drug resistance; MFS, major
survival advantage as well as resulting in low-level
facilitator; MUG, 4-methylumbelliferyl b-D-galactopyranoside; NOR,
resistance to antimicrobial agents that also are substrates
norfloxacin; PENT, pentamidine; PY, pyronin Y; qRT-PCR, quantitativereverse-transcription-PCR; RD, rhodamine 6G; TPP, tetraphenylphos-
for those pumps. Such low-level resistance can have
significant consequences (see below).
Biocide- and dye-induced efflux pump overexpression
Dyes such as acriflavine (AF), ethidium bromide (EB),
associated with the appearance of mepR point mutations
rhodamine 6G (RD) and pyronin Y (PY) have significant
that inactivated this negative regulator. MDR efflux pump
utility in the research laboratory, including nucleic acid
overexpression in S. aureus thus is a relatively common
staining and the study of transport processes in eukaryotic
consequence of biocide and dye exposure.
and microbial cells (Horobin & Kiernan, 2002). Inparticular, EB is a good substrate for many bacterial effluxpumps and susceptibility to it in the presence and absence
of an efflux pump inhibitor has been used as a screen for
Bacterial strains, plasmids, media and reagents. S. aureus
the presence of efflux-related resistance mechanisms
SH1000, which is a derivative of S. aureus NCTC 8325-4 in which
(DeMarco et al., 2007). Structural features of substrates
the rsbU mutation has been repaired, was used as a control strain
of bacterial MDR pumps of the major facilitator (MFS),
(Horsburgh et al., 2002). Eight clinical strains (three meticillin-
small multidrug resistance (SMR), and perhaps the multi-
susceptible and five meticillin-resistant) from a collection of unique
drug and toxin extrusion (MATE) families include
bloodstream isolates of S. aureus used in a study of the frequency of
hydrophobic moieties and a positive charge; many dyes
baseline MDR efflux pump overexpression (DeMarco et al., 2007),each overexpressing no more than one MDR efflux pump, were
and biocides share these characteristics and both groups of
selected at random and employed as test strains (Table 1). Strains and
compounds are commonly transported by an individual
plasmids used for functional analyses of mepR mutants are listed in
MDR pump (Hassan et al., 2007; Poole, 2005).
Table 2. Reagents were the highest grade available and along withmedia were obtained from Sigma or BD Biosciences. Unless otherwise
Examination of the Staphylococcus aureus genome reveals
noted, the incubation temperature for all experiments was 35 uC.
numerous potential MDR efflux-pump-encoding genes (seewww.membranetransport.org). Some of those that have been
Antimicrobial susceptibility testing. MICs were determined in
studied in detail include QacA and QacB, highly similar MFS
duplicate using a microdilution procedure with and without reserpine
pumps that are encoded on plasmids, NorA and MdeA, both
(20 mg ml21) according to Clinical and Laboratory Standards
chromosomally encoded MFS pumps, and MepA, a MATE-
Institute (CLSI) guidelines (CLSI, 2006). Compounds selected forevaluation included AF, benzalkonium chloride (BAC), cetrimide
family MDR pump that also is chromosomally encoded
(CET), chlorhexidine (CHX), crystal violet (CV), dequalinium
(Kaatz et al., 2005a; Paulsen et al., 1996b). More recently
(DEQ), EB, norfloxacin (NOR), pentamidine (PENT), PY, RD, and
described are the NorB, NorC and SdrM MFS pumps, the
tetraphenylphosphonium bromide (TPP). One or more of these
genes for which are also chromosomal (Truong-Bolduc et al.,
compounds have been shown to be substrates for many of the known
2005, 2006; Yamada et al., 2006). Of considerable interest is
S. aureus MDR efflux pumps (Hassan et al., 2007). A reserpine-
the sepA gene, positioned immediately downstream of sdrM,
mediated MIC reduction of at least fourfold was considered indicative
which encodes a structurally unique transporter that has
of efflux of that compound and hereafter will be referred to as asignificant reserpine effect. For production of mutants by the single-
some similarity to the SMR family of proteins (Narui et al.,
exposure approach, MICs were also determined by agar dilution in
2002). Unlike all other S. aureus SMR family pumps
order to incorporate accurate drug concentrations in selection plates.
described to date, SepA is encoded on the chromosome.
Compounds used for single- and multiple-exposure mutant produc-tion were chosen from those showing no significant reserpine effect in
With the exception of SdrM all the transport proteins just
microdilution MIC testing. This approach allowed for simple
described are highly conserved among strains for which
identification of efflux-related phenotypes by the appearance of a
genome sequence data are available (n513; 95–100 %
significant reserpine effect for one or more common pump substrates.
omes/lproks.cgi). SdrM is conserved in 10 of 13 sequenced
Mutant production. Test organisms were exposed to low to
strains (98–100 % homology), but less conservation is
moderate concentrations of various biocides and dyes to determineif such exposure resulted in the appearance of mutants overexpressing
present in MRSA252 (87 %) and in Mu3 and Mu50, both
one or more MDR efflux pumps. For single-exposure mutant
of which have a deletion of residues 206–291. Each of the
production, organisms were grown overnight in Mueller–Hinton
above-described transporters has various biocides and dyes
broth (MHB) and then diluted 100-fold into fresh, pre-warmed
as substrates and some also efflux hydrophilic fluoroquin-
MHB. This culture was incubated with shaking to an OD660 of 0.7, as
olone antimicrobial agents such as norfloxacin, ciproflox-
previous experiments had shown that this represented the late
acin and moxifloxacin. Susceptibility to these clinically
exponential growth phase (data not shown). Cells were recovered by
relevant agents may be sufficiently reduced by MDR pumps
centrifugation and resuspended in 1 ml fresh MHB. Serial dilutionand plating techniques were employed to determine the log
such that the emergence of target-based high-level
(c.f.u. ml21) of this suspension, and aliquots of the undiluted and
resistance is favoured (Markham & Neyfakh, 1996).
serially diluted suspension were plated onto Mueller–Hinton agar(MHA) plates containing two and four times the respective agar
In this study we exposed bloodstream isolates of S. aureus
dilution MIC of the appropriate compound. Compounds used in this
to low to moderate concentrations of several biocides and
procedure included DEQ (strains K3043, K3166, K3221 and K3231),
dyes and examined the resultant mutants for upregulation
CET (strain K3250), BAC (strains SH1000, K3055, K3155, K3221 and
of several MDR efflux pumps, mainly those of chro-
K3250), EB (strains SH1000, K3043, K3155, K3166, K3225 and
mosomal origin. We found that single- and multiple-step
K3231) and RD (strains K3055, K3231 and K3250). Plates were
exposure led to the appearance of mepA, norA, norC and
examined for growth after 48 h incubation.
mdeA-overexpressing mutants, with mepA overexpression
Multiple-exposure mutants were produced using the gradient plate
predominating. Overexpression of mepA most often was
approach (Bryson & Szybalski, 1952). Initial gradients consisted of
zero to two times the agar-dilution MIC of BAC (strains K3055,
K3221 and K3250), EB (strains SH1000, K3043, K3155, K3166, K3225
and K3231), or RD (all strains). After 24 h incubation organisms
from the leading edge of growth were passed again until growthoccurred across the entire plate. This process was repeated using
gradually increasing drug gradients to a maximum of 20, 30 and 3 mg
ml21 for BAC, EB and RD, respectively, and generally was completed
Several colonies from single-exposure plates and the leading edges ofgrowth from the final gradient plates were streaked onto MHA to
recover single colonies of potential mutants. One colony from each
exposure condition was selected at random and microdilution MICs
in the absence and presence of reserpine were determined. These
organisms were passed three additional times on MHA followed by
repeat MIC testing to ensure maintenance of a significant reserpine
effect for at least one test compound compared with the respective
parental strain. Mutants meeting this condition were considered
qRT-PCR. The expression of mepA, norA-B-C, mdeA, sdrM and sepA
in parent and putative MDR pump-overexpressing mutants was
determined using quantitative reverse-transcription PCR (qRT-PCR)
as described previously (DeMarco et al., 2007). Beacon Designer 7.01
(Premier Biosoft International) was used to design Taqman probes
and primers based on the genome sequence of S. aureus 8325, which
were purchased commercially (Operon Biotechnologies). qRT-PCR
was performed in triplicate using the Superscript III Platinum One-
Step kit (Invitrogen) and an ABI 7500 Fast Real-Time PCR system
(Applied Biosystems) with parameters of 45 uC for 30 min, 95 uC for2 min, and 40 cycles of 1 min at 95
and positive controls were included and 16S rRNA was used as the
endogenous control. The comparative threshold cycle method was
used to calculate relative gene expression compared with that of S.
aureus SH1000; expression of each studied gene in this strain was
considered to be 1.0 (Livak & Schmittgen, 2001).
Based on earlier experience with the strain set from which test
organisms used in this study were selected, qacA/B is very infrequent
in Detroit area S. aureus bloodstream isolates (DeMarco et al., 2007). Thus, using primers designed based on the sequence of qacA
(GenBank sequence X56628) that also had complete homology with
qacB, routine PCR was employed to detect the presence or absence of
EB efflux assay. All parent and each stably resistant single- and
multiple-exposure mutant underwent a determination of EB efflux in
real time to demonstrate efflux pump function. The procedure wasperformed using a fluorometric approach as described previously
(Kaatz et al., 2000). Experiments were performed in duplicate and
results were expressed as mean total efflux over a 5 min time-course.
Results for mutant strains were divided by those for the respectiveparent to normalize the data, which for mutants then represented an
n-fold change compared with the parent. An increase of twofold or
greater was considered significant. The effect of reserpine (20 mg
ml21) was also determined and expressed as percentage reduction of
RNA slot blotting. Northern analyses were performed to verify qRT-
PCR results. RNA from strains exhibiting overexpression of pump
genes was immobilized on a nylon membrane using a Bio-Rad Bio-
Dot SF apparatus (Bio-Rad) and methods exactly as described
previously (Ausubel et al., 2005). A PCR-generated fragment of the
appropriate gene served as a probe, and labelling of the probe,
hybridization and detection were performed using the BrightStar
Psoralen-Biotin labelling, NorthernMax and BrightStar BioDetect kitsaccording to the manufacturer’s directions (Ambion). 16S rRNA was
used as the endogenous control and data generated for the
Biocide- and dye-induced efflux pump overexpression
Table 2. Strains and plasmids used for MepR functional analyses
S. aureus vector containing a tetracycline-inducible promoter
controlling expression of cloned genes; Cmr
*Cmr, confers chloramphenicol resistance.
appropriate parent strain were used as the standard to which mutant
were observed for as few as one (CHX for both K3166 and
K3231) to as many as five tested substrates (SH1000). Forthose compounds chosen for use in mutant production,
Sequencing. Promoter and coding regions of MDR pump genes
agar-dilution MICs were within twofold of those deter-
having increased expression relative to parent strains were amplifiedby PCR and then sequenced in both directions using an automated
mined from the microdilution results (data not shown).
method by the Applied Genomics and Technology Center, Wayne
The presence of a significant reserpine effect for at least one
State University (Sanger et al., 1977). For strains overexpressing norA-
compound in all parent strains indicated that some efflux
B-C the sequence of mgrA and its promoter was determined as MgrA
pump activity was probably present at baseline.
is known to affect the expression of each of these genes (Luong et al.,2006). In addition, for strains overexpressing mepA the sequence ofmepR, which encodes the repressor of mepA expression and is
encoded immediately upstream of mepA, was determined also. DSGene 1.5 (Accelrys) was used for nucleotide sequence analyses. The
With the exception of K3225 all parent strains were
sequence of the respective parental strain was the standard with which
exposed to at least two single-exposure selecting agents,
and putative mutants were recovered in all cases except fortwo- and fourfold MIC BAC exposures for strain K3250
MepR functional analyses. Point mutations in mepR resulting in
and fourfold MIC RD exposures for all strains. Single-
MepR amino acid substitutions were observed in two mepA-
exposure mutants appeared at frequencies ranging from 1
overexpressing mutants (see Table 4). The mepR coding region fromeach of these mutants was amplified using PCR and cloned into
in 106 to 1 in 109 c.f.u. at twofold MIC exposures and 1 in
pALC2073, producing pK580 and pK582 (Table 2). Strains containing
107 to 1 in 1010 c.f.u. at fourfold MIC exposures. Serial
pACL2073-based constructs were grown in the absence and presence
passage of putative mutants on drug-free media identified
of 50 ng tetracycline ml21 to assess the effect of induction of plasmid-
instability of the resistance phenotype for 9 of 17 single-
MepR is a repressor of both mepR and mepA expression. This
increases in the remaining eight mutants are listed in
property was exploited to assess the functional integrity of mutant
Table 3. MIC increases ranged from as little as twofold to as
MepR proteins using a fluorescent b-galactosidase assay as described
high as 32-fold. Interestingly, fourfold or greater MIC
previously, employing 4-methylumbelliferyl b-D-galactopyranoside
increases were infrequently observed for the selecting agent
(MUG) as a substrate (Kaatz et al., 2006). b-Galactosidase activity was
(two out of eight mutants). For example, exposure of
expressed in MUG units; 1 MUG unit51 pmol MUG cleaved per
strains K3043, K3166 and K3221 to DEQ resulted in
minute per OD600 unit. Experiments were performed in triplicate,and the expression of chromosomal mepR in SA-K2916-R, SA-K2916-
fourfold or greater MIC increases for eight, five and seven
R (Q18P), and SA-K2916-R (G97E) was quantified by integrating the
substrates, respectively, but these substrates did not include
area beneath the expression curves with SigmaPlot 10.0 (Systat
The multiple-exposure, or gradient plate, process resultedin the recovery of putative mutants in all cases in which
this procedure was applied. As for single-exposuremutants, not all strains maintained their resistance profilesafter serial passage in drug-free media. However, the
proportion that did was greater than observed for single-
MICs for parent strains with and without reserpine are
exposure organisms (14 out of 18 versus 8 out of 17).
given in Table 1. Fourfold or greater decreases in MICs
Instability was observed for SH1000 (EB exposure), K3055
Table 3. Susceptibility changes for single- and multiple-exposure mutants (compared with parent strain)
*Selection conditions and compound. SE and ME, single- and multiple-exposure. See Table 1 footnote for testcompound abbreviations.
and K3250 (BAC exposure), and K3155 (RD exposure).
increased expression of any pump gene with the exception
Fourfold or greater MIC increases for selecting agents were
of K3055, K3225 and K3250, which had elevated norA
not always observed but were more common than in
expression at baseline compared with SH1000 (6.6-, 7.1-,
single-exposure mutants (11 out of 14; 79 %).
Increased expression of mepA predominated (8 out of 22
mutants, or 36 %), resulting most commonly after multiple
All parental strains and stable mutants were analysed for
exposure [BAC (1), EB (3), RD (2)] but also occurring with
their expression levels of mepA, norA-B-C, mdeA, sepA and
single exposure [DEQ (1) and EB (1)]. Increased norA
sdrM compared with that of SH1000. Data for mutants
expression was observed in three mutants (14 %) and
then were normalized to those of the appropriate parent,
followed single exposure to DEQ (1) and multiple exposure
and a fourfold or greater increase in expression compared
to EB (2). Increased mdeA and norC expression were
with the parent was considered significant (Table 4). qRT-
observed only after multiple-exposure to RD (one mutant
PCR results for genes demonstrating significantly increased
each). One multiple-exposure mutant overexpressed two
expression were verified by slot-blot analyses (data not
pump genes (mepA and norA; K3155 ME-EB). No
shown). qacA/B were found to be absent from this strain
increased expression of norB, sepA or sdrM was observed.
set using a PCR-based screening approach (data not
The magnitude of increased pump gene expression ranged
shown). None of the parental strains demonstrated
from as little as 7-fold to more than 450-fold. There also
Biocide- and dye-induced efflux pump overexpression
Table 4. Gene expression, EB efflux, and sequence analyses
Premature mepR stop after 10 codons; A302S
*Strains with increased norA expression at baseline are indicated (qnorA). DSelection conditions and compound. SE and ME, single- and multiple-exposure. See Table 1 footnote for test compound abbreviations. dCompared with parent strain. §Normalized to parent strain. ||Only differences from parent strain are shown. ND, Not determined.
were five single- and five multiple-exposure mutants in
which no change in pump gene expression was identified.
The 10 mutants not overexpressing any studied pump gene
Like the qRT-PCR data, EB efflux data for mutants were
were omitted from this analysis. For the remaining 12
normalized to the respective parent strain (Table 4).
mutants sequence data are provided in Table 4; only
Interestingly, one mutant having a ninefold increase in
differences from the respective parent strain are shown. For
norC expression had no change in EB efflux (K3225 ME-
strains overexpressing mepA a variety of mutations were
RD). Similar behaviour with respect to EB efflux was
observed in mepR, including four strains having changes
evident in 8 of the 10 mutants for which no increased
resulting in the creation of premature stop codons very
expression of the genes studied here was identified. Where
early in the reading frame and two others with point
increased EB efflux was observed, reserpine generally was
mutations resulting in amino acid substitutions in MepR.
effective in reducing the efflux by at least 50 % (data not
The mepRA sequence of the remaining two mepA-
overexpressing strains was unchanged from that of the
respective parent. A novel point mutation resulting in an
A302S MepA substitution was observed for one mutant.
MDR efflux pumps are an important mechanism by which
The single mdeA-overexpressing mutant was found to have
bacteria can evade the effect(s) of antimicrobial agents. This
a point mutation that changed the 235 motif from
resistance mechanism has received considerable attention in
GTGCTA to TTGCTA as well as a GAT transversion at
recent years, and there are ongoing efforts to develop
the +2 position of mdeA mRNA (Huang et al., 2004). The
inhibitors of MDR pumps. Effective inhibitors would
235 motif change results in a closer match to the
expand the antibacterial armamentarium to include pump
substrates and reduce the likelihood of the emergence ofhigh-level
Three parental strains overexpressed norA and in each case
Unfortunately, inhibitors evaluated to date have not been
a previously described mutation associated with increased
broad-spectrum in their activity. Inhibitors of MFS pumps
norA expression, consisting of an insertion of AAT
are not active against resistance-nodulation-division (RND)
immediately 39 to the 210 promoter motif, was observed
family pumps such as AcrB of Escherichia coli or MexB of
(DeMarco et al., 2007). Overexpression of norA in two
Pseudomonas aeruginosa, organisms of considerable medical
mutants (K3155 ME-EB and K3231 ME-EB) was associated
importance. Likewise, RND inhibitors do not have activity
with a novel potential promoter up-mutation, consisting of
against MFS pumps (Kaatz, 2005). Nevertheless, safe and
an AAG transition between the 235 and 210 motifs. This
effective inhibitors of the major classes of bacterial MDR
alteration disturbs an inverted repeat that has been shown
pumps, even if separate compounds are required for MFS
to be important for the full repressive effect of MgrA
and RND inhibition, would be a welcome addition to
(Kaatz et al., 2005b). The sequence of mgrA for both of
these strains was wild-type. The sequences of norA andmgrA were wild-type in the remaining norA-overexpressing
Previous studies in our laboratory have shown that
mutant (K3043 SE-DEQ). Likewise, the sequences of mgrA
exposure of S. aureus to low concentrations of fluoroquin-
and norC were unchanged from those of the parent strain
olone antimicrobial agents can result in upregulation of
for the norC-overexpressing mutant (K3225 ME-RD).
NorA and MepA, but the effect of biocide exposure ondrug pump expression in this organism has not beenevaluated in detail (Kaatz & Seo, 1995; Kaatz et al., 2000).
Others have shown previously that exposure of E. coli to
The induction of wild-type mepR expression in SA-K2916-
antiseptic agents such as pine oil and triclosan results in the
R resulted in an 83 % reduction in chromosomal mepR
upregulation of the AcrB MDR pump, which includes
expression, whereas minimal to no effect was observed for
several antimicrobial agents in its substrate profile (Levy,
strains SA-K2916-R (Q18P) and SA-K2916-R (G97E),
2002; McMurry et al., 1998; Moken et al., 1997). Our data
now extend this observation to S. aureus in that we have
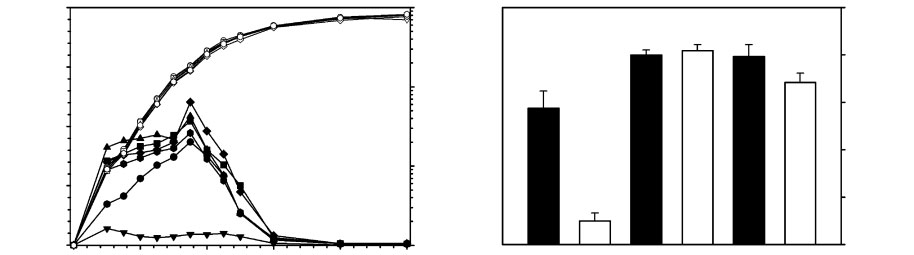
Fig. 1. Expression of chromosomal mepR in SA-K2916 containing pK434 (SA-K2916-R), pK580 [SA-K2916-R (Q18P)], andpK582 [SA-K2916-R (G97E)]. (a) mepR expression as a function of growth: $ and ., SA-K2916-R; & and X, SA-K2916-R(Q18P); m and {, SA-K2916-R (G97E). In each case data represented by the first and second symbols were obtained in theabsence and presence of 50 ng tetracycline ml”1, respectively. Open symbols illustrate growth of test organisms. Error bars areomitted for the sake of clarity. (b) Cumulative expression of mepR over the entire experiment. Black and white bars illustrateexpression in the absence and presence of tetracycline, respectively. 1 and 2, SA-K2916-R; 3 and 4, SA-K2916-R (Q18P); 5and 6, SA-K2916-R (G97E). Means±SD are plotted.
Biocide- and dye-induced efflux pump overexpression
shown that exposure of clinical isolates to low concentra-
lation. Differences in sensitivity between the qRT-PCR and
tions of a variety of biocides and dyes can produce mutants
EB efflux assays also may have played a role.
with increased expression of one or more MDR pumps.
Ten mutants demonstrated significant MIC increases but
Even though the increases in biocide MICs observed in
no change in expression of any of the pumps included in
these mutants were moderate, such organisms are likely to
our study. It is possible that these MIC changes may be the
have a survival advantage in environmental niches where
result of a non-pump-related mechanism(s). However, it is
more likely that these organisms have increased expression
Compounds chosen for mutant production were those
of pumps other than those we evaluated as all had a
showing no more than a twofold MIC reduction for
significant reserpine effect for at least one of the tested
parental strains in the presence of the efflux pump
substrates. EB efflux of 8 of these 10 strains, including the
inhibitor reserpine. This approach resulted in variable
RD multiple-exposure mutants of K3043, K3055, K3231
agents being employed for individual test strains. However,
and K3250, the RD and DEQ single-exposure mutants of
the purpose of our work was not to compare one agent
K3055 and K3221, respectively, and EB single- and
with another with respect to the frequency of emergence of
mutants but rather to determine, in general, if biocides and
increased from that of the appropriate parent strain
dyes commonly employed in the hospital or laboratory
(Table 4). Five of these eight mutants were derived from
environments could induce overexpression of MDR efflux
parental strains that overexpressed norA; EB efflux in those
parental strains was ¢47 %. The baseline activity of NorA
The absence of stably raised MICs for the selecting agent
in mutants derived from these parents almost certainly
employed for some mutants was of considerable interest.
would obscure any contribution to EB efflux from other
This observation was more common for single-exposure
pumps. It is also conceivable that EB may be a poor
than multiple-exposure mutants (6 out of 8 versus 3 out of
substrate or not a substrate at all for these as-yet-
14, respectively). The lower incidence of this characteristic
unidentified pumps. Baseline increased norA expression
in multiple-exposure mutants most likely relates to the
and the fact that EB is a relatively poor NorC substrate
increased opportunities for accumulation of mutations in
probably accounts for the lack of change in EB efflux by the
strains repeatedly exposed to a particular compound.
RD multiple-exposure mutant of K3225, which over-
Additive or synergistic effects on susceptibility are expected
expressed norC (Hassan et al., 2007).
as the number of resistance-conferring mutations increases.
Among the genes included in our screen, overexpression of
However, for those mutants demonstrating instability of
mepA predominated (8 out of 22 mutants). Mutations in
selecting-agent MICs, substrate affinity for the pump(s) in
the gene encoding the MepR repressor were responsible in
question may play a role. MIC changes for compounds
six cases, including four having premature truncations of
with lower affinity are likely to be less pronounced than for
MepR and two with amino acid substitutions (Q18P and
those that are better substrates. Alternatively, it is
G97E) resulting in severe impairment in MepR activity
conceivable that unidentified mutations may be present
(Fig. 1). With respect to the two mutants having no
in these mutants affecting susceptibility to selecting agents
mutations in the mepRA region, one or more mutations in
in a direction opposite to that of the MDR pumps. It also
must be recalled that we employed a definition of a stable
Overexpression of mepA has been observed in another
fourfold or greater MIC increase as significant, but if
strain in which the mepRA sequence is wild-type (Kaatz et
twofold or greater were employed then the selecting agent
al., 2005a). It is apparent that mepA expression can be
would be included in all cases (data not shown).
Instability of the resistance phenotype to serial passage in
In addition to augmented mepA expression, an A302S
the absence of drug was intriguing but not necessarily
MepA substitution was identified in K3043 ME-EB. It
surprising. Pump overexpression in the absence of
seems unlikely that this change contributed to the 12-fold
substrate could confer a selective disadvantage by way of
increase in EB efflux observed for this mutant. Single
wasted resources or perturbed growth kinetics. Reversion
amino acid differences, both within transmembrane helices
to a wild-type phenotype in the absence of selective
and in extramembrane loops, are known to alter substrate
pressure would reverse these potential disadvantages.
specificity, as has been shown for QacA/B and the
The magnitude of pump gene overexpression did not
tetracycline efflux transporter TetA (Paulsen et al., 1996a;
necessarily correlate with increases in EB efflux observed.
Sapunaric & Levy, 2005). In addition, there are conserved
This discrepancy is exemplified by the BAC and RD
charged residues in the extramembrane loop between
multiple-exposure mutants of K3221, where mepA expres-
transmembrane helices 2 and 3 in many MFS efflux
sion was 12- and 450-fold increased, respectively, with
proteins, changes in which can adversely affect protein
similar increases in EB efflux (Table 4). Transcript quantity
function (Yamaguchi et al., 1992). However, to the best of
may not correlate with translated protein. There may be a
our knowledge, mutations resulting in increased efficiency
maximal amount of MDR pump tolerated and in this
of transport for a particular substrate have not been
situation excess transcripts are degraded prior to trans-
identified. Introduction of the A302S mutation into MepA
and a comparison of the kinetics of EB efflux with that of
DeMarco, C. E., Cushing, L. A., Frempong-Manso, E., Seo, S. M.,
wild-type MepA would be required to establish or refute
Jaravaza, T. A. & Kaatz, G. W. (2007). Efflux-related resistance to
Staphylococcus aureus. Antimicrob Agents Chemother 51, 3235–3239.
Two of the three mutants overexpressing norA were found to
Hassan, K. A., Skurray, R. A. & Brown, M. H. (2007). Active export
have the same mutation in the promoter region, consisting
proteins mediating drug resistance in staphylococci. J Mol Microbiol
of an AAG transition between the 235 and 210 motifs.
This change disrupts an inverted repeat, but whether or not
Horobin, R. W. & Kiernan, J. A. (2002). Conn’s Biological Stains, 10th
it affects norA expression requires further study. Likewise,
edn. Oxford, UK: BIOS Scientific Publishers.
the mdeA-overexpressing mutant had a mutation resulting
Horsburgh, M. J., Aish, J. L., White, I. J., Shaw, L., Lithgow, J. K. &
in the conversion of the 235 motif into a sequence that
Foster, S. J. (2002). sB modulates virulence determinant expression
more closely matches the preferred consensus sequence and
and stress resistance: characterization of a functional rsbU strainderived from Staphylococcus aureus 8325-4. J Bacteriol 184, 5457–
may improve promoter strength (Szoke et al., 1987). It also
had a GAT transversion at the +2 position that could affect
Huang, J., O’Toole, P. W., Shen, W., Amrine-Madsen, H., Jiang, X.,
mRNA stability, but further work would be required to
Lobo, N., Palmer, L. M., Voelker, L., Fan, F. & other authors (2004).
assess this possibility formally. Increased norA and norC
Novel chromosomally encoded multidrug efflux transporter MdeA in
expression not associated with sequence changes in known
Staphylococcus aureus. Antimicrob Agents Chemother 48, 909–917.
regulatory elements, such as the promoter regions or known
Kaatz, G. W. (2005). Bacterial efflux pump inhibition. Curr Opin
regulatory loci such as mgrA, is likely to result from
mutational alterations in as-yet-uncharacterized loci that
Kaatz, G. W. & Seo, S. M. (1995). Inducible NorA-mediated
affect the expression of these genes.
multidrug resistance in Staphylococcus aureus. Antimicrob AgentsChemother 39, 2650–2655.
Exposures to MDR pump substrates, whether thosesubstrates are clinically relevant antimicrobial agents or
Kaatz, G. W., Seo, S. M., O’Brien, L., Wahiduzzaman, M. & Foster, T. J. (2000). Evidence for the existence of a multidrug efflux transporter
compounds used to disinfect the hospital environment, can
distinct from NorA in Staphylococcus aureus. Antimicrob Agents
result in the emergence of S. aureus strains adapted to the
presence of these compounds. Acquisition of such strains
Kaatz, G. W., McAleese, F. & Seo, S. M. (2005a). Multidrug resistance
by patients may compromise the therapy of infections
in Staphylococcus aureus due to overexpression of a novel multidrug
caused by them. Prudent use of antimicrobial agents and
and toxin extrusion (MATE) transport protein. Antimicrob Agents
the use of biocides that are not known to be MDR pump
substrates may reduce the frequency at which MDR-pump-
Kaatz, G. W., Thyagarajan, R. V. & Seo, S. M. (2005b). Effect of
overexpressing strains are found. Alternatively, the com-
promoter region mutations and mgrA overexpression on transcrip-
bination of a pump inhibitor with an antimicrobial agent
tion of norA, which encodes a Staphylococcus aureus multidrug efflux
or biocide will reduce the emergence of such strains and
transporter. Antimicrob Agents Chemother 49, 161–169.
Kaatz, G. W., DeMarco, C. E. & Seo, S. M. (2006). MepR, a repressorof the Staphylococcus aureus MATE family multidrug efflux pumpMepA, is a substrate-responsive regulatory protein. Antimicrob AgentsChemother 50, 1276–1281.
Levy, S. B. (2002). Active efflux, a common mechanism for biocideand antibiotic resistance. J Appl Microbiol 92 (Suppl.), 65S–71S.
This work was supported by VA Research Funds.
Livak, K. J. & Schmittgen, T. D. (2001). Analysis of relative geneexpression data using real-time quantitative PCR and the 22DDC(T)
Luong, T. T., Dunman, P. M., Murphy, E., Projan, S. J. & Lee, C. Y.
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman,
(2006). Transcription profiling of the mgrA regulon in Staphylococcus
J. G., Smith, J. A. & Struhl, K. (2005). Current Protocols in Molecular
aureus. J Bacteriol 188, 1899–1910.
Markham, P. N. & Neyfakh, A. A. (1996). Inhibition of the multidrug
Bateman, B. T., Donegan, N. P., Jarry, T. M., Palma, M. & Cheung,
transporter NorA prevents emergence of norfloxacin resistance in
A. L. (2001). Evaluation of a tetracycline-inducible promoter in
Staphylococcus aureus. Antimicrob Agents Chemother 40, 2673–2674.
Staphylococcus aureus in vitro and in vivo and its application in
McBain, A. J., Rickard, A. H. & Gilbert, P. (2002). Possible implications
demonstrating the role of sigB in microcolony formation. Infect
of biocide accumulation in the environment on the prevalence of
bacterial antibiotic resistance. J Ind Microbiol Biotechnol 29, 326–330.
Boyce, J. M. (2007). Environmental contamination makes an
McDonnell, G. & Russell, A. D. (1999). Antiseptics and disinfectants:
important contribution to hospital infection. J Hosp Infect 65
activity, action, and resistance. Clin Microbiol Rev 12, 147–179.
McMurry, L. M., Oethinger, M. & Levy, S. B. (1998). Overexpression of
Bryson, V. & Szybalski, W. (1952). Microbial selection. Science 115,
marA, soxS, or acrAB produces resistance to triclosan in laboratory
and clinical strains of Escherichia coli. FEMS Microbiol Lett 166,
CLSI (2006). Approved standard M7-A7. Methods for dilution
antimicrobial susceptibility tests for bacteria that grow aerobically,
Moken, M. C., McMurry, L. M. & Levy, S. B. (1997). Selection of
7th edn. Wayne, PA: Clinical and Laboratory Standards Institute.
multiple-antibiotic-resistant (Mar) mutants of Escherichia coli by
Biocide- and dye-induced efflux pump overexpression
using the disinfectant pine oil: roles of the mar and acrAB loci.
Smith, K., Gemmell, C. G. & Hunter, I. S. (2008). The association
Antimicrob Agents Chemother 41, 2770–2772.
between biocide tolerance and the presence or absence of qac genes
Narui, K., Noguchi, N., Wakasugi, K. & Sasatsu, M. (2002). Cloning
among hospital-acquired and community-acquired MRSA isolates. J
and characterization of a novel chromosomal drug efflux gene in
Staphylococcus aureus. Biol Pharm Bull 25, 1533–1536.
Szoke, P. A., Allen, T. L. & deHaseth, P. L. (1987). Promoterrecognition by Escherichia coli RNA polymerase: effects of base
Paulsen, I. T., Brown, M. H., Littlejohn, T. G., Mitchell, B. A. & Skurray,R. A. (1996a). Multidrug resistance proteins QacA and QacB from
substitutions in the 210 and 235 regions. Biochemistry 26, 6188–6194.
Staphylococcus aureus: membrane topology and identification of
Truong-Bolduc, Q. C., Dunman, P. M., Strahilevitz, J., Projan, S. J. &
residues involved in substrate specificity. Proc Natl Acad Sci U S A 93,
Hooper, D. C. (2005). MgrA is a multiple regulator of two new efflux
pumps in Staphylococcus aureus. J Bacteriol 187, 2395–2405.
Paulsen, I. T., Brown, M. H. & Skurray, R. A. (1996b). Proton-
Truong-Bolduc, Q. C., Strahilevitz, J. & Hooper, D. C. (2006). NorC, a
dependent multidrug efflux systems. Microbiol Rev 60, 575–608.
new efflux pump regulated by MgrA of Staphylococcus aureus. Antimicrob Agents Chemother 50, 1104–1107.
Poole, K. (2005). Efflux-mediated antimicrobial resistance. J AntimicrobChemother 56, 20–51.
Yamada, Y., Hideka, K., Shiota, S., Kuroda, T. & Tsuchiya, T. (2006). Gene cloning and characterization of SdrM, a chromosomally-
Russell, A. D. (2003). Biocide use and antibiotic resistance: the
encoded multidrug efflux pump, from Staphylococcus aureus. Biol
relevance of laboratory findings to clinical and environmental
situations. Lancet Infect Dis 3, 794–803.
Yamaguchi, A., Nakatani, M. & Sawai, T. (1992). Aspartic acid-66 is
Sanger, F., Nicklen, S. & Coulson, A. R. (1977). DNA sequencing with
the only essential negatively charged residue in the putative
chain-terminating inhibitors. Proc Natl Acad Sci U S A 74, 5463–5467.
hydrophilic loop region of the metal-tetracycline/H+ antiporter
Sapunaric, F. M. & Levy, S. B. (2005). Substitutions in the
encoded by transposon Tn10 of Escherichia coli. Biochemistry 31,
interdomain loop of the Tn10 TetA efflux transporter alter
tetracycline resistance and substrate specificity. Microbiology 151,2315–2322.
NEW ORDER FORM - OTC DRUGS (Order online at www.weeksandleo.com ) Minimum order quantity 12 Items with * min order quantity 6 Description National Brand Packaging Retail Price Total New COLDS ~ SINUS ~ ALLERGY COUGH ~ COLD SLEEP AID ACID REDUCER (H2 BLOCKER) Private Label - OTC Line (G:\Document Control\Pricing\2009) NEW ORDER FORM - OTC DRUGS (Orde
Saft Interview - Article Aircraft Technology Engineering & Maintenance 1) Saft’s expertise ranges across a number of different industries. What are the key characteristics of aviation batteries and what special requirements must they fulfil? Aviation batteries are a crucial link in aircraft safety. With 80+ years of experience in aviation, one Saft battery takes off every two seconds, S
 respective parent. A novel point mutation resulting in an
A302S MepA substitution was observed for one mutant.
respective parent. A novel point mutation resulting in an
A302S MepA substitution was observed for one mutant.