Microsoft word - synthetische evolutionstheorie.doc
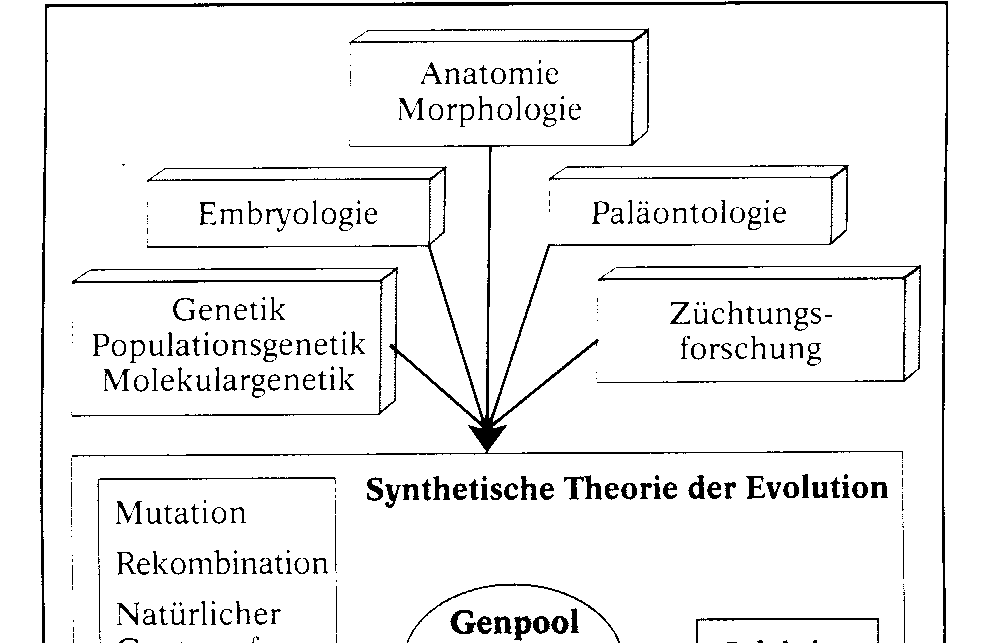
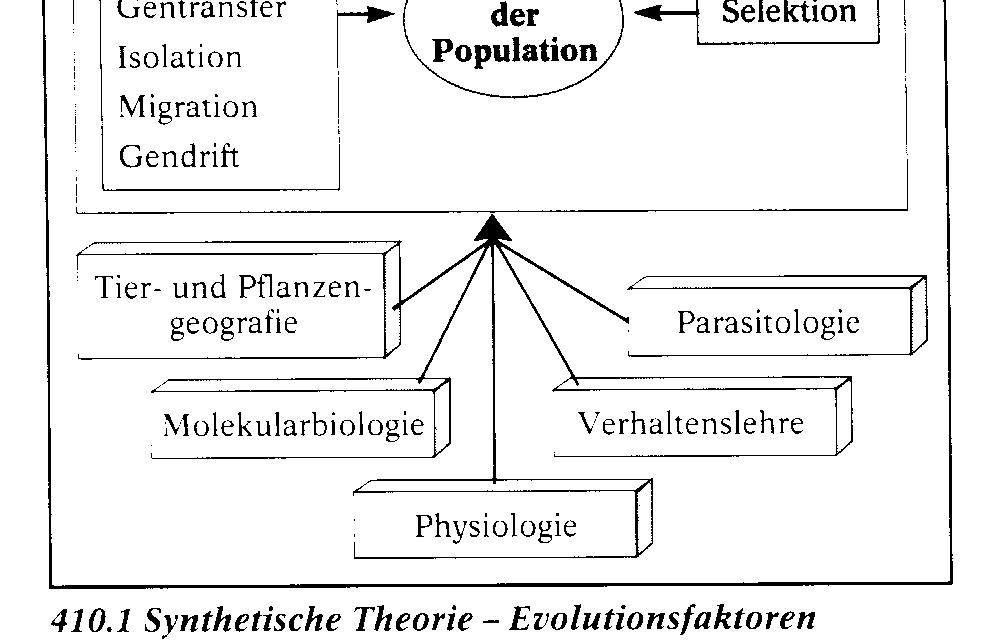
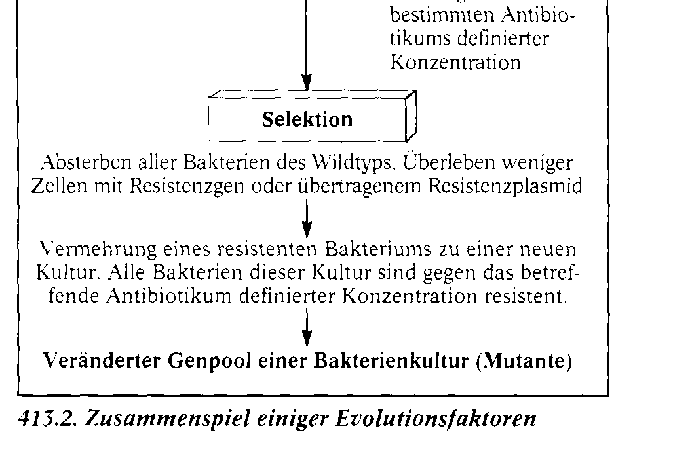
Die synthetische Theorie der Evolution. Diese Theorie fasst die Erkenntnisse aller biologischen Teilwissenschaften zusammen. Sie berû¥ck- sichtigt die grundlegenden Gesichtspunkte DARWINS, also die ûberproduktion an Nachkommen, die erblichen Variationen (Mutationen) und die Selektion. Sie wird aber durch eine Reihe weiterer wichtiger Evolutionsfaktoren ergûÊnzt und gilt nach heutigem Erkenntnisstand als am besten be- grû¥ndet. WûÊhrend der Evolution wird der Genpool, das ist die Gesamtheit der in einer Population vorhandenen Gene, verûÊndert. Am Beispiel einer Bakterienkultur lûÊsst sich das Zusammenspiel der Evolutionsfaktoren und damit ein kleines Stû¥ck Bakterien-Evolution verfolgen. Zû¥chtet man das Bakterium Staphylococcus au- reus in Petrischalen auf reinen Agar-NûÊhrbûÑden und auf NûÊhrbûÑden mit dem Antibiotikum Penicil- lin, so sind die Schalen mit reinem Agar-NûÊhrboden spûÊter voller Bakterienkolonien, wûÊhrend die Schalen mit Penicillin nur wenige Kolonien enthalten. Sie haben sich aus Bakterien entwickelt, die vom Penicillin nicht abgetûÑtet wurden. Sie sind gegenû¥ber diesem Antibiotikum in der gewûÊhlten Konzentration resistent. Hier liegt nicht etwa eine Anpassung im Sinne von Ge- wûÑhnung an Penicillin vor. WiderstandsfûÊ- higkeit gegen Penicillin ist keine erworbene, an die Nachkommen weitergegebene Eigen- schaft, vielmehr haben diese Bakterien Gene, die fû¥r die WiderstandsfûÊhigkeit verantwort- lich sind. Wie kommt es dazu? Gene von Bakterien mutieren. Unter zehn Millionen Zellen befindet sich im Durchschnitt eine, bei der ein ãResistenzgenã vorhanden ist. Auch die ûbertragung von mutierten Genen von einem Bakterium auf ein anderes (Re- kombination) kommt vor. Rekombination und Mutation sind ungerich- tet. Sie verûÊndern in unserem Beispiel den Genpool der Bakterienpopulation. Sie stellen das Material, an dem die Selektion ansetzt, fû¥r den Ausleseprozess bereit. Als Selekti- onsfaktor wirkt hier das Penicillin. Es tûÑtet die Bakterien des Wildtyps ab und selektio- niert die wenigen Zellen mit einem Resistenz-Gen heraus. Alle aus diesen resistenten Bakterien gewonnenen neuen Bakterienkulturen bestehen aus Bakterien mit der neuen Eigenschaft. Im gewûÊhlten Beispiel wirken die drei Evolutionsfaktoren Mutation, Rekombination und Selektion zusammen. Weitere, den Genpool beeinflussende Evolutionsfaktoren sind: die Wanderung von In- dividuen aus einer Population in eine andere (Migration) und die Isolation, bei der Teile der Popu- lation von anderen Teilen isoliert werden. Hinzu kommen Gendrift und natû¥rlicher Gentransfer.
Mutation, Rekombination, natû¥rlicher Gentransfer
Mutation. Bakterien teilen sich unter optimalen Kulturbedingungen alle 20 Minuten. In 24 Stunden kûÑnnen also theoretisch 272 = 4.7 x 1021 Zellen entstehen. Ist unter Hunderttausenden von Bakterien im KûÑrper eines erkrankten Menschen ein einzelnes infolge einer Mutation gegen ein bestimmtes An- tibiotikum resistent, so kann es den Einsatz dieser medizinischen Waffe û¥berleben. Es wird also aus einer riesigen Zahl von Erregern, die alle abgetûÑtet werden, ausgelesen. Seine Resistenz gibt es an alle Nachkommen weiter. Damit ist das eingesetzte Medikament wirkungslos geworden. Die HûÊufigkeit, mit der ein einzelnes Gen mutiert, heiût Mutationsrate. Sie liegt fû¥r Bakterien im Durchschnitt bei 10-7 . Das bedeutet, dass ein einzelnes Gen in nur einer unter 10 Millionen Zellen mutiert. Die Mutationsrate ist niedrig. Infolge der unvorstellbar hohen Nachkommenzahl der Bakte- rien ist die Wahrscheinlichkeit fû¥r das Auftreten einer Mutation aber doch relativ hoch. Bei vielzelligen Organismen kûÑnnen Mutationen in den KûÑrperzellen oder in den Keimzellen auf- treten. Nur, wenn sie in Ei- oder Spermazellen bzw. deren Vorstadien vorliegen, kûÑnnen bei ge- schlechtlicher Fortpflanzung Mutanten entstehen. Bei vielzelligen Organismen schûÊtzt man die Mu- tationsrate fû¥r ein einzelnes Gen auf 10-6 . Unter einer Million Keimzellen tritt also nur eine auf, die am betreffenden Genort eine Mutation aufweist. Die Zahl der Gene eines eukaryotischen Orga- nismus ist aber sehr hoch. Man schûÊtzt sie je nach Art auf 104 - bis 106. Ein Organismus mit 105 Genen hûÊtte bei einer Mutationsrate von 10-6 also immerhin in 1ã10 seiner Keimzellen eine Mu- tation aufzuweisen. Trotz niedriger Mutationsrate ist infolge der hohen Genzahl die Wahrschein- lichkeit fû¥r das Auftreten einer Mutation relativ hoch. Man nimmt z. B. an, dass 10% bis 40% aller Keimzellen des Menschen ein mutiertes Gen tragen. Mutationen verûÊndern den Genbestand oder Genpool einer Population qualitativ. Sie sorgen fû¥r den unerschûÑpflichen Vorrat an genetischer Va- riation in einer Population. Sie stellen das Material fû¥r den Evolutionsprozess bereit. Damit sind sie ein wesentlicher Evolutionsfaktor Rekombination. Durch die freie Kombination der Chromosomen bei der Keimzellbildung wird eine sehr groûe Zahl genetisch unterschiedlicher Gameten ausgebildet. Durch Chromosomenstû¥ck-Austausch wûÊhrend der Meiose wird diese Zahl noch erhûÑht. Bei der Verschmelzung von Ei- und Spermazelle entstehen immer wieder neue Genotypen. Dem 3. MENDELschen Gesetz entsprechend findet eine Neukom- bination der Gene und damit auch der Merkmale statt. Diese Neukombination von Erbanlagen bei der sexuellen Fortpflanzung nennt man Rekombination. Durch die verschiedenen Prozesse der Re- kombination entsteht in einer Population genetische Vielfalt. Rekombinationen sind fû¥r die Entste- hung neuer Genotypen noch wichtiger als die Mutationen. GûÊbe es plûÑtzlich keine Mutationen mehr, entstû¥nden trotzdem noch in Hunderten von Generationen stûÊndig neue Genotypen durch Re- kombination. Die Zahl der KombinationsmûÑglichkeiten der Gene innerhalb einer Population ist un- vorstellbar hoch. Deshalb kûÑnnen bei weitem nicht alle theoretisch mûÑglichen Kombinationen in einer Population tatsûÊchlich vertreten sein. Natû¥rlicher Gentransfer. Austausch von genetischer Information erfolgt nicht nur zwischen verschiedenen Bakterienarten. Es gibt auch Genû¥bertragung von Bakterien auf hûÑhere Pflanzen. So befûÊllt Agrobacterium tumefa- ciens verwundete Pflanzen und transferiert dabei ein Stû¥ck Plasmid-DNA in den Kern der Pflan- zenzelle. Die eingefû¥hrten Gene zwingen die Pflanze zu abnormem Wachstum. Es bilden sich Tu- more, die Wurzelhalsgallen. Die Plasmid Gene zwingen die Pflanze auûerdem, AminosûÊuren, die ihr fremd sind, zu erzeugen. Sie werden an den Wurzeln freigesetzt und dienen der ErnûÊhrung der Bakterien. Das Bakterium baut also eigene genetische Information in das Wirtsgenom ein und lûÊsst sich gewissermaûen ãnach eigenem Rezept seine Suppe kochenã. Genaustausch scheint auch zwi- schen nicht-verwandten Vielzellern mûÑglich. So nimmt man an, dass bei Hû¥lsenfrû¥chtlern gefunde- ne HûÊmoglobin-Gene tierischen Ursprungs sind. Es ist also denkbar. dass neue Organismen nicht NF BIOLOGIE 13.2 EVOLUTION
nur durch Mutation, sondern auch durch den Erwerb neuer Genfolgen von anderen Lebewesen ent-standen sind.
Hybridisierung als Evolutionsfaktor? 2500 untersuchte BûÊlge von Darwinfinken und Freilandbeobachtungen haben gezeigt, dass es bei diesen VûÑgeln Hybridisierung gibt. Es wurden gemischte Paare mit lebensfûÊhigen Nachkommen und Rû¥ckkreuzungen der Hybriden mit einer Ursprungsart beobachtet. Nach neuesten Studien wurden bei jeder zehnten Vogelart Hybridisierungen beobachtet. Auch bei vielen anderen Tierarten scheint der Austausch genetischer Information zwischen Elternpopulationen eine Rolle fû¥r ihre Entwicklung zu spielen. Hybridisierung ist damit als weiterer Evolutionsfaktor zu diskutieren.
EvolutionsûÑkologie
Selektion. Fû¥r den EvolutionsûÑkologen ist die natû¥rliche Selektion ein fundamentales Prinzip der ûkologie. Ein Beispiel verdeutlicht das: Der extrem kalte und lange Winter 1946/47 in Mitteleuropa fû¥hrte zur Verminderung der Zahl der Insekten. Dadurch litten die Maulwû¥rfe unter Nahrungsmangel, der ei- nen Selektionsdruck auf sie ausû¥bte. Genetisch bedingt kleinere Tiere hatten einen Selektionsvor- teil, sie kamen mit weniger Nahrung aus. GrûÑûere Tiere verhungerten eher. Dadurch war der pro- zentuale Anteil grûÑûerer Tiere nach diesem Winter gesunken, Das zeigt uns: Die Auslese setzt am PhûÊnotyp an, Genotypische Unterschiede. die phûÊnotypisch nicht in Erscheinung treten, werden von der Selektion nicht erfasst. Die Evolutionsfaktoren Mutation, Rekombination und Gentransfer stellen fû¥r die natû¥rliche Ausle- se, die Selektion, das Material bereit. Diese gibt dann dem Evolutionsprozess eine Richtung, indem sie die weniger tauglichen Individuen zurû¥ckdrûÊngt, wûÊhrend sie die anderen Individuen der glei- chen Population begû¥nstigt. Die Maulwû¥rfe in unserem Beispiel wurden den Umweltbedingungen ãangepasstã. Angepasstheit steht in strengem Umweltbezug. Bei der Entscheidung, ob ein Merkmal in einer Population positi- ven oder negativen Selektionswert hat, muss man die jeweiligen Umweltbedingungen berû¥cksichti- gen. Unter extremem Nahrungsmangel war die geringe KûÑrpergrûÑûe fû¥r die Maulwû¥rfe vorteilhaft. Unter anderen Bedingungen, z. B. bei Zunahme der natû¥rlichen Feinde, kûÑnnte die Kleinheit der Maulwû¥rfe wegen geringerer VerteidigungsfûÊhigkeit einen negativen Selektionswert haben. Die Angepasstheit eines Lebewesens kann zu einem bestimmten Zeitpunkt an einem bestimmten Ort hoch, an anderem Ort, zu anderer Zeit dagegen gering sein.
Das Zusammenwirken von Mutation und Selektion kann man an einer Bakterienkultur von E.coli beobachten. (Abb. links) Impft man E.coli (Wildtyp) definierter Konzentration auf ei- nen VollnûÊhrboden 1, so erhûÊlt man nach Bebrû¥tung einen dichten Bakterienrasen. Im Parallelversuch unter Antibioti- kumzugabe entstehen dagegen nur wenige Kolonien 2. Das Antibiotikum wirkt als Selektionsfaktor. In beiden FûÊllen entstehen nur wenige Mutationen. Man spricht deshalb von einem schwachen Mutationsdruck. Auf dem VollnûÊhrboden ist auch der Selektionsdruck schwach. Das Antibiotikum sorgt dagegen fû¥r einen starken Selektionsdruck, sodass nur einzelne (im Beispiel 3)Kolonien aus resistenten Bakterien hervorgehen. Die anderen werden abgetûÑtet. Wiederholt man den Versuch nun mit Coli-Bakterien, die mit
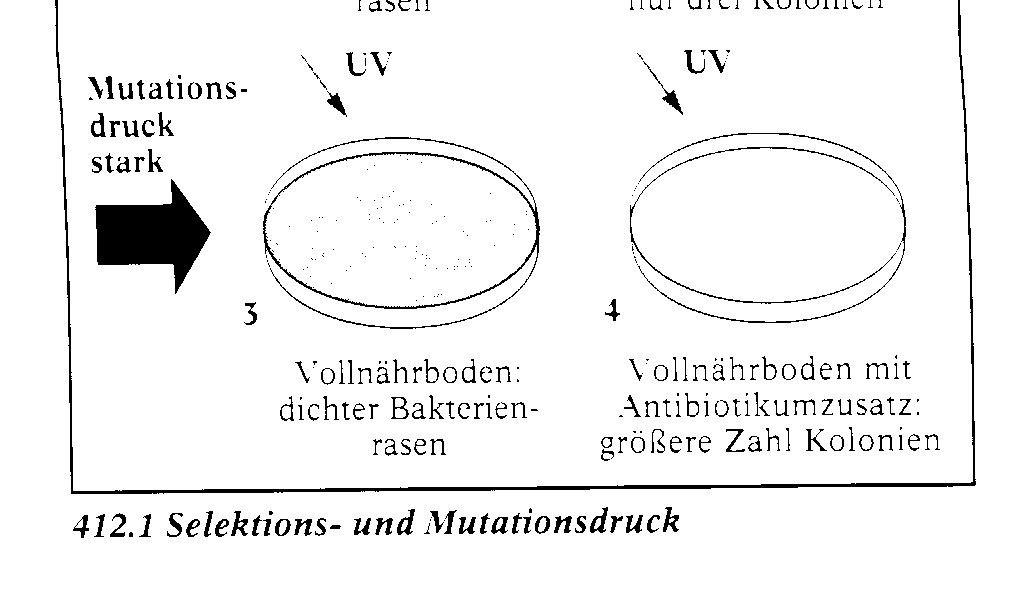
UV-Licht bestrahlt wurden. so erhûÊlt man auf dem VollnûÊhr-
boden unverûÊndert einen dichten Bakterienrasen . Auf dem Antibiotikum-NûÊhrboden findet man diesmal erheblich mehr Kolonien als im Fall der unbestrahlten Bakterien 4. Dem starken Selek- tionsdruck wirkt in diesem Fall durch die erhûÑhte Mutationsrate ein starker Mutationsdruck entge- gen. Das Antibiotikum wirkt also in einer Kultur von Escherichia coli als Selektionsfaktor. Es û¥bt auf die Bakterien einen Druck aus, der zum Verschwinden der nicht resistenten Zellen fû¥hrt. Die- sem Selektionsdruck wirkt der Mutationsdruck entgegen, der durch die in den Kulturen entstehen- den Mutanten hervorgerufen wird. Als Selektionsfaktoren kûÑnnen alle Einflû¥sse der unbelebten Natur, wie Niederschlag, Temperatur, WindverhûÊltnisse, Bodenbeschaffenheit und chemische Bedingungen, wirken. Das gilt auch fû¥r Or- ganismen. Sie kûÑnnen als Konkurrenten um Nahrung und Wohnraum auftreten. Parasiten und Krankheitserreger spielen eine Rolle. Auch die geschlechtliche Auslese bei der Partnerwahl ist von Bedeutung. Selektion bewirkt, dass bestimmte Individuen mehr Nachkommen haben und damit mehr zum Gen- pool der Folgegeneration beitragen als andere. Diesen grûÑûeren Erfolg bei der Fortpflanzung wer- den diejenigen haben, die bei der Nahrungskonkurrenz und bei der Partnerwahl erfolgreicher sind, bessere Brutpflege betreiben, schlechte Umweltbedingungen besser ertragen kûÑnnen usw. Beispiele fû¥r das Wirken der Selektion. Zu Zeiten der Industrialisierung war die Rinde von BûÊumen in Industriegebieten durch Ruû schwarz gefûÊrbt. Dunkle Mutanten von Insekten waren dort vor Fressfeinden besser geschû¥tzt. Sie hatten gewissermaûen eine Tarntracht und setzten sich deshalb gegenû¥ber ihren heller gefûÊrbten Konkurrenten durch. Wegen des dunklen Farbstoffs Melanin in Flû¥geln und KûÑrper hat man den Begriff Industriemelanismus geprûÊgt. Die dunkle Mutante des Birkenspanners ist dafû¥r ein Beispiel. Tarnung durch einen verblû¥ffend umgestalteten KûÑrper zeigen z. B. die Stabheuschrecken, die von den ûsten der Pflanzen. auf denen sie leben, nicht zu unterscheiden sind. Beim wandelnden Blatt, einem Heuschrecken-Verwandten, sind die Flû¥gel, die Flû¥geldecken und die Beine so gestaltet. dass das Tier einem Blatt zum Verwechseln ûÊhnelt. Wespen sind auffûÊllig schwarz-gelb gefûÊrbt. Sie signalisieren mit der Warnfarbe ihren Feinden: Vorsicht, Gefahr! Giftige oder unbekûÑmmliche Tie- re werden hûÊufig von nichtgiftigen im Aussehen ãnachgeahmtã. Diese ãvorgetûÊuschte" Warntracht wirkt als schû¥tzende ûhnlichkeit und heiût Mimikry. Auf Inseln findet man Insekten mit reduzierten oder gar fehlenden Flû¥geln. Sie haben offenbar ei- nen Selektionsvorteil, weil flugunfûÊhige Insekten nicht vom Wind aufs Meer hinausgeweht werden. Manche HûÑhlentiere sind blind. Dieser Mangel wirkt sich nicht negativ aus, weil Blindheit bei der Lebensweise im Dunkeln keinen negativen Auslesewert hat.
Der Industriemelanismus zeigt, dass der Selektionsdruck durch Fressfeinde beim Birkenspanner relativ schnell zur Verwandlung und damit zum Ange- passtsein an die dunkle Baumrinde fû¥hrt. Ein weiteres Beispiel ist die Strandkrabbe. An der Ostkû¥ste Nordamerikas drang sie seit 1900 in be- stimmte Regionen vor. Eines ihrer Beutetiere ist eine Schnecke. Vergleicht man GehûÊuse dieser Schneckenart aus der Zeit vor 1900 mit GehûÊusen aus spûÊterer Zeit, so stellt man fest, die Tiere haben einen flacheren, dickwandi- geren GehûÊusetyp als vor dem Einwandern der Strandkrabbe. Die dick- wandige Form ist fû¥r die Krabben schwerer zu knacken. In krabbenfreien Gebieten haben die Schnecken noch immer ihre hûÑheren. dû¥nnwandigen GehûÊuse. Hier zeigt sich erneut, wie wirksamer Selektionsdruck zu einem relativ schnellen Wandel fû¥hrt. Isolation
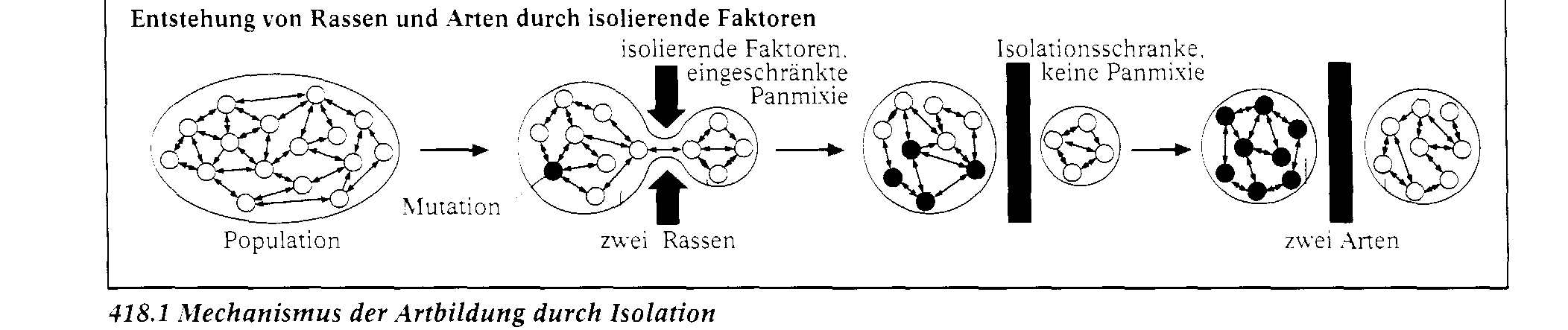
Gemsen lebten vor der Eiszeit in den meisten Hochgebirgen Europas. Heute leben Gemsen nur noch in einzelnen, getrennten Regionen. Nach der Eiszeit wurden die Tiere aus ihrem geschlosse- nen Wohngebiet hierher zurû¥ckgedrûÊngt. Durch die geografische Trennung ist es nun mûÑglich. dass sich in zukû¥nftigen, sehr langen ZeitrûÊumen aus ihnen vielleicht verschiedene Gemsenarten entwi- ckeln werden. Eine neue Eigenschaft entsteht immer durch Mutationen. Wenn sie nicht nachteilig sind, verbreiten sie sich û¥ber die gesamte Population. Prinzipiell kann sich jedes Individuum einer Population zusammen mit jedem andersgeschlechtlichen derselben Art fortpflanzen. TatsûÊchlich geschieht das nicht, aber û¥ber lûÊngere Zeit hinweg tritt eine Durchmischung des Erbguts ein. Dies bezeichnet man als Panmixie. Alle Faktoren, die diese beeintrûÊchtigen, werden als Iso- lationsfaktoren bezeichnet. Geografische Isolation. Die Beuteltiere Australiens sind das klassische Beispiel fû¥r geografische Isolation. Als sie entstan- den, hatte sich der australische Kontinent schon von den anderen Kontinenten getrennt, sodass kei- ne MûÑglichkeit des genetischen Austauschs mit anderen Formen mehr bestand. Andere FûÊlle finden wir auf neu entstehenden Inseln, die besiedelt werden. Auf den GalûÀpagosinseln entstanden auf die- se Weise die Darwin-Finken, die einmal vom sû¥damerikanischen Festland einwanderten und dann hier eine vûÑllig eigenstûÊndige Entwicklung erlebten. Ethologische Isolation. Bei den Regenpfeifern gibt es zwei Arten. die sich ûÊuûerlich recht wenig, dafû¥r aber in ihrem Ver- halten stark unterscheiden, Die Abbildung zeigt zwei Verhaltensweisen, die das belegen. Das Im- ponierverhalten des Sandregenpfeifers unterscheidet sich stark von dem des Flussregenpfeifers. Da
die Verhaltensweisen bei den Artgenossen angeborene AuslûÑsemechanismen ansprechen, reagiert die eine Art nicht auf das Imponierverhalten der anderen und umge-kehrt. Einen starken Bezug zur Vermehrung und damit zur Weitergabe der Gene hat das AblûÑseverhalten am Nest. Beim Brû¥ten lûÑsen sich Sandregenpfeifer ab, indem der ankommende Vogel den brû¥tenden Partner von der Seite her von den Eiern drûÊngt. Das AblûÑsen der Flussre-genpfeifer erfolgt dagegen so, dass der ankommende Vogel von hinten auf das Nest fliegt und der brû¥tende dieses nach vorn verlûÊsst. Verhaltensunterschiede wie diese kûÑnnen dazu beitragen. dass eine Kommunikation zwischen mûÑglichen Kreuzungspartnern so stark er-schwert ist, dass keine Kreuzung mehr stattfindet.
Jahreszeitliche Isolation. Wasserfrosch und Grasfrosch sind zwar genetisch kreuzbar, ihre Balzphasen liegen aber zu unter- schiedlichen Zeiten im Jahr. Deshalb kommt es normalerweise in der Natur nicht zu Kreuzungen. ûkologische Isolation. Unter den Darwin-Finken gibt es solche mit spitzen SchnûÊbeln, die besonders gut Insekten fangen kûÑnnen, und solche, die mit massiven KernbeiûerschnûÊbeln KûÑrner und Samen aufknacken. Durch neue Erbmerkmale erhalten Lebewesen hûÊufig die MûÑglichkeit. neue ErnûÊhrungs- und WohnmûÑg- lichkeiten zu nutzen. Dadurch sind sie dem Konkurrenzdruck entzogen und kûÑnnen sich nahezu un- gestûÑrt entwickeln. Adaptive Radiation. Wenn in der Evolution durch Anpassung unterschiedliche Entwicklungslinien entstehen, spricht man von adaptiver Radiation. Die Darwin-Finken z. B. spalteten sich durch Adaptation (Anpas- sung) an verschiedene LebensrûÊume in mehrere Arten auf. Auch der Pferdestammbaum zeigt im MiozûÊn und im PliozûÊn adaptive Radiationserscheinungen . Die Aufspaltung in verschiedene Ra- dien (Strahlen) erfolgt unmerklich und langsam. Anfangs sind die Unterschiede zwischen den Individuen noch so gering, dass sie sich noch miteinander paaren und fruchtbare Nachkommen haben kûÑnnen. In diesem Fall spricht man von Rassen. Die Rassen der Menschen sind dafû¥r ein Beispiel. Treten im Laufe der Entwicklung von Lebewesen jedoch mehr trennende Eigenschaften auf, so finden kaum noch sexuelle Kontakte statt. Durch zû¥chterische AktivitûÊten kûÑnnen noch Nachkommen erzeugt werden, die dann aber meist steril sind. Maultiere und Maulesel z. B., die aus der Kreuzung von Pferd und Esel entstehen, sind unfruchtbar. Wenn die Unterschiede so groû geworden sind, dass keinerlei sexuelle Kontakte mehr stattfinden und auch keine fortpflanzungsfûÊhigen Nachkommen mehr entstehen, spricht man von zwei getrennten Arten.
GrundlûÊggande virologi, 5 p Ht 2006 HûÊlsouniversitetet LinkûÑping Examinationsuppgift: Skriftlig redovisning av Influensa A Sammanfattning: Influensa A ûÊr en luftburen och mycket smittsam virusinfektion som drabbar framfûÑr allt de ûÑvre delarna av luftvûÊgarna hos dûÊggdjur och fûËglar. Influensa- virus kan ge ûËterkommande vûÊrldsomspûÊnnande epidemier. Enligt WHO:s statistik (2005) dû
Submission to the Standing Committee on Community Services Bill 31: Mental Health Amendment Act I'm very concerned about the "unintentional consequences" of Bill 31. In terms of my special interest area, the collateral damage is very likely going to include an increase of both chemical and physical restraints, instead of adequate and appropriate care, for elderly persons whose dem
 Die synthetische Theorie der Evolution.
Die synthetische Theorie der Evolution. 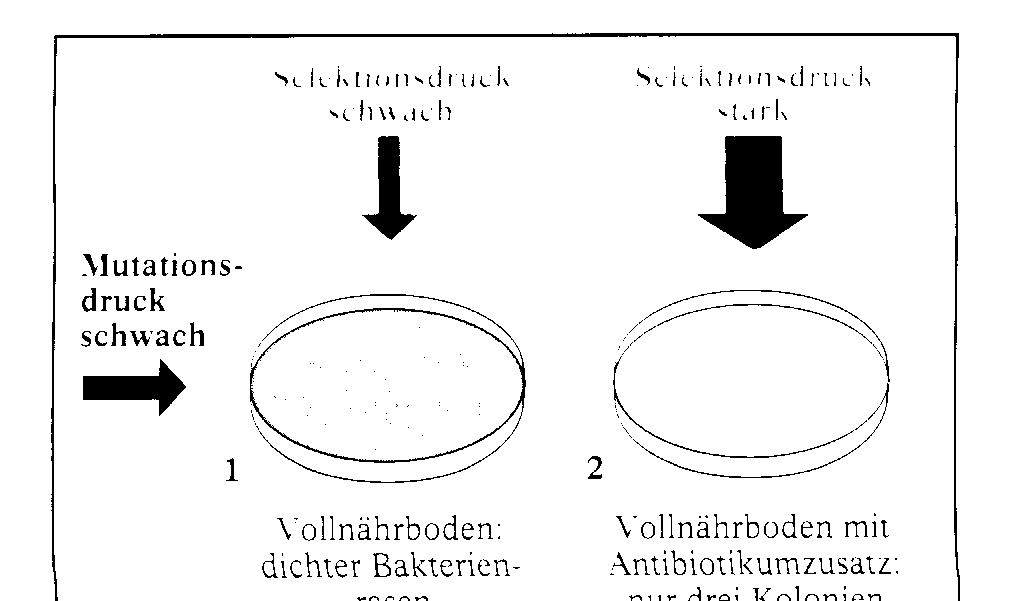
 nur durch Mutation, sondern auch durch den Erwerb neuer Genfolgen von anderen Lebewesen ent-standen sind.
Hybridisierung als Evolutionsfaktor?
nur durch Mutation, sondern auch durch den Erwerb neuer Genfolgen von anderen Lebewesen ent-standen sind.
Hybridisierung als Evolutionsfaktor? 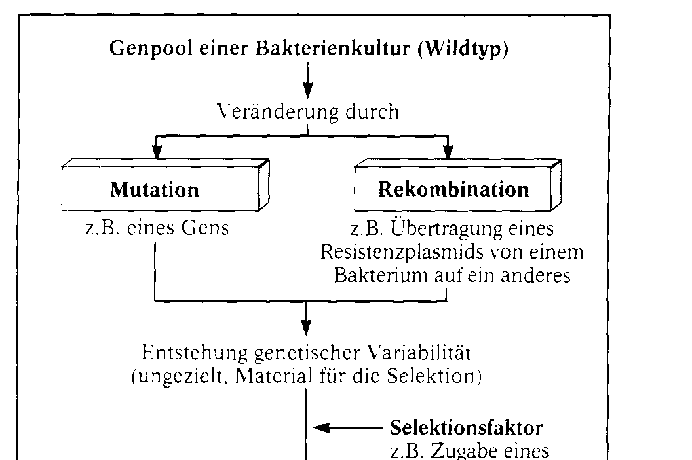
 boden unverûÊndert einen dichten Bakterienrasen . Auf dem Antibiotikum-NûÊhrboden findet man
boden unverûÊndert einen dichten Bakterienrasen . Auf dem Antibiotikum-NûÊhrboden findet man 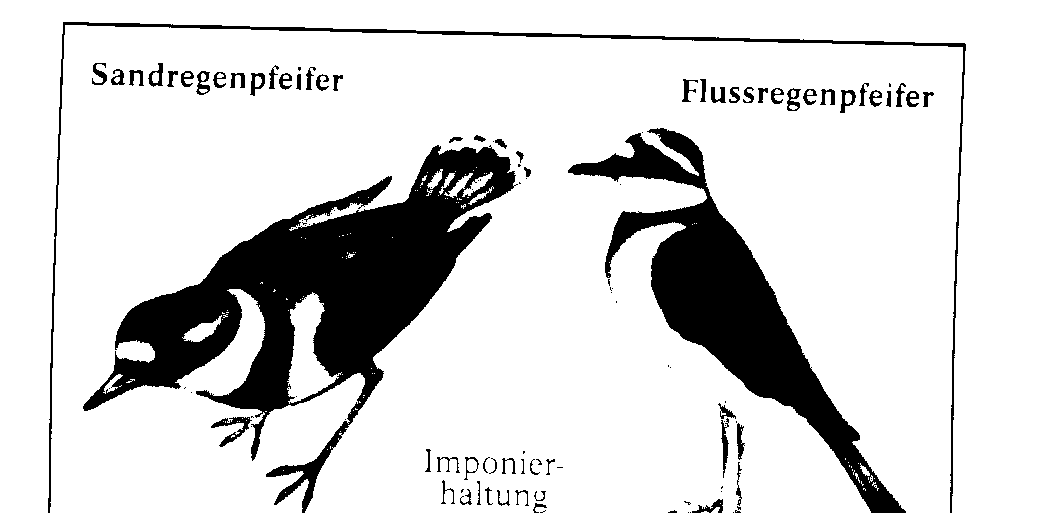
 Isolation
Isolation 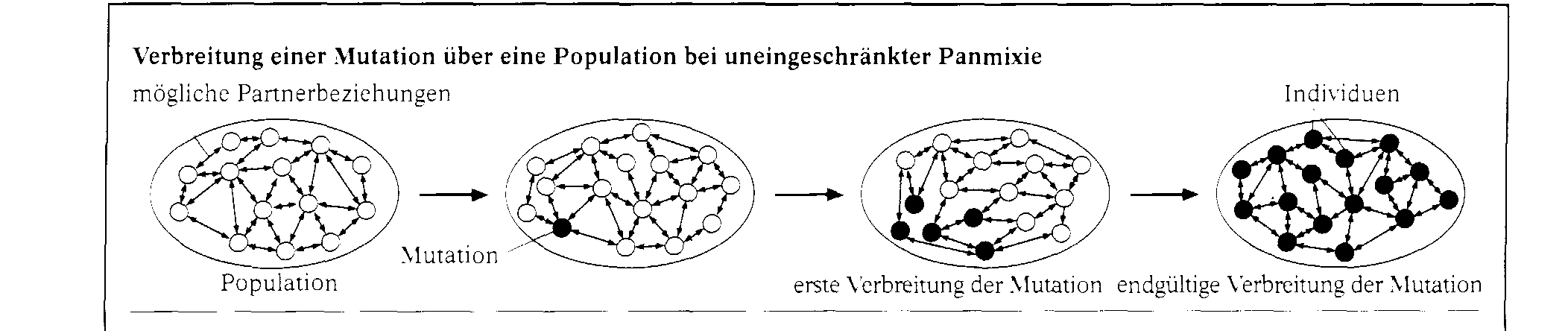
 Adaptive Radiation.
Adaptive Radiation.