Cyclooxygenase mechanisms Lawrence J Marnett
Several advances have occurred in the past year in our
polyunsaturated fatty acid autoxidation (Figure 2) [1]. The
understanding of cyclooxygenase catalysis. The role of specific
13-pro(S)-hydrogen is removed and O2 traps the incipient
heme oxidation states in the formation of catalytically
pentadienyl radical at C-11. The 11-peroxyl radical
competent tyrosyl radicals has been defined; the identity of
cyclizes at C-9 and the carbon-centered radical generated
physiological hydroperoxide activators has been established;
at C-8 cyclizes at C-12, producing the endoperoxide. The
and the participation of individual amino acids in substrate
allylic radical generated is trapped by O2 at C-15 to form
binding and oxygenation has been elucidated.
the 15-(S)-peroxyl radical; this radical is then reduced toPGG2. Several pieces of experimental evidence support
Addresses
this mechanism: firstly, a significant kinetic isotope effect
Departments of Biochemistry and Chemistry, Vanderbilt University
is observed for the removal of the 13-pro(S)-hydrogen [2];
School of Medicine, Nashville, Tennessee 37232, USA;
secondly, carbon-centered radicals are trapped during
e-mail: marnett@toxicology.mc.vanderbilt.edu
catalysis [3]; and thirdly, minor oxidation products are
Current Opinion in Chemical Biology 2000, 4:545–552
formed that arise by oxygen trapping of an allylic radicalintermediate at positions 13 and 15 [4,5]. A variation of the
mechanism in Figure 2 in which the 13-pro(S)-hydrogen is
2000 Elsevier Science Ltd. All rights reserved.
removed as a proton and the incipient carbanion is oxi-
Abbreviations
dized to a radical is theoretically possible. However,
oxygenation of 10,10-difluoroarachidonic acid to
11-(S)-hydroxyeicosa-5,8,12,14-tetraenoic acid is inconsis-tent with the occurrence of a carbanion intermediate
Introduction
because the latter would rapidly eliminate fluoride to form
Cyclooxygenases (COXs) catalyze the committed step in
a conjugated diene [6]. The absence of endoperoxide-con-
the conversion of arachidonic acid to prostaglandins and
taining products derived from 10,10-difluoroarachidonic
thromboxane. They oxygenate arachidonic acid to the
acid has been suggested to indicate the importance of a
hydroperoxy endoperoxide PGG2 (prostaglandin G2),
C-10 carbocation in PGG2 synthesis [7]. However, the pro-
followed by reduction of PGG2 to the alcohol PGH2
posed cationic mechanism postulates that endoperoxide
(Figure 1). PGH2 is converted by isomerases to
formation precedes removal of the 13-pro(S)-hydrogen [7].
prostaglandins and thromboxane, which exert numerous
This is inconsistent with the results of isotopic labeling
physiological and pathophysiological effects. Thus, COX
experiments of arachidonic acid oxygenation [2].
enzymes play a key role in the biosynthesis of a family ofimportant bioactive lipids. But it is the interesting chem-
Identity of the protein oxidant
istry which they catalyze that is the focus of this review.
The oxidant that removes the 13-pro(S) hydrogen
Recent advances in the mechanism of arachidonic acid oxy-
appears to be a tyrosyl radical derived from Tyr385
genation, the identity of the protein oxidant, the pathway
(Figure 2) [8]. This residue is interposed between the
of enzyme activation, and the nature of enzyme−substrate
heme prosthetic group and the cyclooxygenase active
interactions will be described. Amino acid designations are
site and is ideally positioned to interact with a bound
given based on the COX-1 numbering system.
fatty acid molecule [9,10,11••]. Transient tyrosyl radicalsare detected during cyclooxygenase catalysis and they
Mechanism of arachidonate oxygenation
oxidize arachidonic acid to carbon-centered radicals
The conversion of arachidonic acid to PGG2 can be formu-
[12••]. It has been difficult to assign the identity of the
lated as a series of radical reactions analogous to those of
tyrosyl radicals based solely on electron paramagnetic
The conversion of arachidonic acid to PGH2. COX catalyzes the oxidation of arachidonic
physiological and pathophysiological effects.
R1 = CH2CH=CH(CH2)3CO2H R2 = C5H11 AH2 = Reducing substrate
Mechanisms
Overall mechanism of COX activation and catalysis. A hydroperoxide
Tyr385 to a tyrosyl radical (upper half of figure). The tyrosyl radical then
oxidizes the heme prosthetic group to a ferryl-oxo derivative that can be
oxidizes the 13-pro(S) hydrogen of arachidonic acid to initiate the
reduced in the first step of the peroxidase catalytic cycle or can oxidize
cyclooxygenase catalytic cycle (lower half of figure).
resonance (EPR) spectroscopy [13,14]. Multiple hyper-
cyclooxygenase activity but does not eliminate radical
fine splitting patterns are observed that arise from
production following reaction with a hydroperoxide (see
rotational isomers of tyrosyl radicals and possibly tyrosyl
below) [17]. Because there are several tyrosine residues
radicals derived from different amino acids [15,16•]. Site-
at distances from the heme that are comparable to that of
directed mutation of Tyr385 to Phe abolishes
Tyr385, another tyrosine residue may be oxidized when
Cyclooxygenase mechanisms Marnett 547
Tyr385 is absent. Interestingly, the Tyr385Phe mutant
enzyme does not oxidize arachidonic acid to carbon-centered radicals, even though it does produce tyrosyl
radicals following treatment with a hydroperoxide [12••].
The enzymatically generated tyrosyl radical has beentrapped by carrying out reactions of arachidonic acid and
COX-1 in the presence of NO donors [18]. NO quenches
the tyrosyl radical signals, presumably by forming a
nitrosocyclohexadienone. The nitrosocyclohexadienoneis oxidized to an iminoxyl radical and ultimately to
Trapping of the enzymatically generated tyrosyl radical by carrying out
nitrotyrosine (Figure 3). Tryptic digestion and peptide
reactions of arachidonic acid and COX-1 in the presence of NO
mapping reveal the presence of a single nitrated peptide
donors. NO quenches the tyrosyl radical signals, presumably by
that contains a nitrotyrosine at the position in the
forming a nitrosocyclohexadienone. The nitrosocyclohexadienone is
sequence corresponding to Tyr385 [19••]. Formation of
oxidized to an iminoxyl radical and ultimately to nitrotyrosine.
this nitrated peptide requires cyclooxygenase turnover inthe presence of NO and is blocked by the cyclooxygenaseinhibitor indomethacin.
reconstitute the peroxidase activity by providing a distalbase to facilitate proton transfer during Compound I
Role of the heme
The Tyr385 tyrosyl radical is not present in resting enzymeso it must be generated in order to initiate cyclooxygenase
Generation of Compound I by reaction of ferric enzyme
catalysis. Reaction of fatty acid hydroperoxides or organic
with hydroperoxide establishes a thermodynamically
hydroperoxides with the heme prosthetic group generates
favorable sequence of reactions to initiate cyclooxyge-
a higher oxidation state of the heme that oxidizes Tyr385
nase catalysis. The redox potential for Compound I is
(Figure 2) [20]. The higher oxidation state that oxidizes
estimated to be ~1 V by comparison with the analogous
Tyr385 is the ferryl-oxo complex, which is the first inter-
ferryl-oxo complex of horseradish peroxidase [26].
mediate in peroxidase catalysis (Compound I) [20,21].
Reaction of Compound I with a tyrosine residue is
Decay of the visible absorbance of Compound I coincides
with production of the tyrosyl radical [22•].
tyrosyl radical with the doubly allylic hydrogens of apolyunsaturated fatty acid (Eo = 0.6 V) [27,28]. In con-
Alterations in enzyme activity that reduce peroxidase
trast, the redox potentials for the resting ferric enzymes
activity introduce a lag phase in cyclooxygenase activation
are −167 mV and −156 mV for COX-1 and COX-2,
[23]. For example, mutations of the proximal histidine
respectively, making direct oxidation of Tyr385 by ferric
residue to tyrosine (His388Tyr) or the distal histidine to
enzyme highly unfavorable thermodynamically
alanine (His207Ala) reduce peroxidase activity by 2 to
[24••,29]. The low redox potentials of COX-1 and
4 orders of magnitude and induce lag phases of 1 to 2 min-
COX-2 are consistent with the observation that the rest-
utes for attainment of maximal cyclooxygenase activity
ing enzymes are isolated in the ferric form and do not
following addition of arachidonic acid [21,24••]. This lag
contain a spectroscopically detectable tyrosyl radical.
phase is eliminated by addition of exogenous hydroperox-
Theoretically, it is possible that the endothermic nature
ides. The ability of a hydroperoxide to eliminate the lag
of the oxidation of Tyr385 by ferric enzyme is circum-
phase correlates to its ability to serve as a peroxidase sub-
vented by electron tunneling [30]. This may explain the
strate [24••,25]. In the case of the distal histidine mutant
activation of a derivative of COX-1 modified with bro-
(His207Ala), the lag phase also can be eliminated by
moacetamido-indomethacin, which has no detectable
adding large amounts of 2-methylimidazole to chemically
For the distal histidine mutant (His207Ala), thelag phase can be eliminated by adding largeamounts of 2-methylimidazole to chemicallyreconstitute the peroxidase activity. The 2-
facilitate proton transfer during Compound I
initiate cyclooxygenase catalysis (see Figure 2). Mechanisms
biosynthesis with COX activation mediated byperoxynitrite. Hollow arrows representstimulation of gene expression and enzyme
NO synthase PGH synthase Hydroperoxide activators
superoxide dismutase mimetic agents, which prevents
The activation of resting enzyme following addition of
peroxynitrite formation, reduces prostaglandin biosynthe-
arachidonic acid in vitro is due to the presence of trace
sis by up to 85% [42]. An attractive feature of the
amounts of hydroperoxide in the fatty acid preparation.
involvement of peroxynitrite as an activator of cyclooxy-
Activation is completely inhibited by addition of high con-
genase in inflammatory cells is the fact that both the
centrations of glutathione peroxidase and glutathione,
inducible form of nitric oxide synthase and COX-2 are
which reduces fatty acid hydroperoxides [33–35]. Once the
immediate-early genes that are induced by many of the
Tyr385 radical is generated, each enzyme molecule cat-
same agonists and with very similar time courses [44,45].
alyzes several hundred cycles of arachidonic acid
This provides a regulated pathway for the generation of a
oxygenation. Although the tyrosyl radical is reduced to tyro-
hydroperoxide activator coincident with COX-2 expres-
sine when it oxidizes arachidonic acid, the radical is
sion (Figure 5). In fact, prostaglandin synthesis by
regenerated in the last step of each catalytic cycle by oxi-
activated macrophages from iNOS-knockout mice is sig-
dation by the peroxyl radical precursor to PGG2. There is
nificantly reduced compared with synthesis by activated
occasional leakage of the peroxyl radical from the cyclooxy-
macrophages isolated from wild-type mice [46•].
genase active site, which leaves the enzyme in acatalytically inactive form containing fully covalent Tyr385. Enzyme−substrate interactions
Reactivation of cyclooxygenase activity requires reaction of
Considerable attention has focused recently on the bind-
the heme prosthetic group with another molecule of
ing of fatty acid substrates in the cyclooxygenase active
hydroperoxide. This explains the need for the continued
site. The chemical mandates of the synthesis of a bicyclic
presence of hydroperoxide in cyclooxygenase–arachidonic-
peroxide with trans-dialkyl substitution require that the
acid reactions [36]. However, by and large, the
fatty acid be bound in an extended conformation with a
cyclooxygenase catalytic cycle proceeds independently of
sharp bend around carbons 10−13 [47]. Modeling this con-
the peroxidase catalytic cycle once Tyr385 is oxidized to a
formation of arachidonate into the cyclooxygenase active
tyrosyl radical. This is supported by three pieces of evi-
site with the carboxylate ion-paired to Arg120 and the
dence: the ability to isolate PGG2 as the major product of
13-pro(S) hydrogen adjacent to Tyr385 places the ω-end of
arachidonic acid oxygenation [37,38]; detailed kinetic
the fatty acid in a hydrophobic pocket near the top of the
analyses consistent with independent turnover of the per-
active site (Figure 6; [48••]). A conserved glycine residue
oxidase and cyclooxygenase activities after activation [39];
(Gly533) is located close to the end of the fatty acid.
and the ability of site-directed mutants with low peroxidase
Mutation of Gly533 to Ala reduces cyclooxygenase activity
activity to achieve near wild-type cyclooxygenase activity
with arachidonate as substrate by 85% and mutation to Val
once the lag phase is eliminated [24••].
completely eliminates activity. However, both mutantsexhibit undiminished cyclooxygenase activity toward
The identity of the ‘physiological’ hydroperoxide activa-
unsaturated fatty acids containing fewer carbons at their
tor is uncertain, but several possibilities exist. Several
ω-end (e.g. α-linolenic acid, stearidonic acid; [48••]).
different fatty acid hydroperoxides react with COX togenerate Compound I, so lipid hydroperoxides are likely
Confirmation of the importance of the top channel in
activators [40,41•]. Peroxynitrite, the coupling product of
substrate binding is provided by crystal structures of com-
NO and superoxide anion, is also an efficient substrate for
plexes of arachidonate bound to COX-1 reconstituted
the peroxidase of both COX-1 and COX-2 [42]. It acti-
with Co3+–heme (W Smith, personal communication) and
vates the cyclooxygenase activity of either enzyme in the
of PGH2 bound to apoCOX-2 [11••]. In addition, exami-
presence of very high concentrations of glutathione per-
nation of these structures reveals numerous
oxidase and glutathione, and activates COX-1 in intact
protein–fatty-acid interactions, suggesting an active role
smooth muscle cells [43]. Treatment of lipopolysaccha-
for the protein in controlling the regiochemistry and
ride-activated macrophages with membrane-permeant
stereochemistry of arachidonate oxygenation. Of particular
Cyclooxygenase mechanisms Marnett 549
importance is the region around Tyr385 and Trp387. As
stated above, the tyrosyl radical derivative of Tyr385 oxi-dizes the 13-pro(S) hydrogen of arachidonate and, asexpected, it is positioned close to C-13 in both structures. In addition, Trp387 is close to the endoperoxide group inthe COX-2–PGH2 crystal structure, suggesting that itmay restrict the conformation of the 11-peroxyl radical tofacilitate cyclization at C-9 [11••]. Indeed, mutation of
Trp387 to Phe or Tyr does not abolish oxygenase activitybut reduces the yield of PGH2 20-fold [11••,49]. Crystallography and site-directed mutagenesis also suggestthat the protein controls the stereochemistry of O2 addi-tion to radicals at C-11 and C-15 by steric hindrance
([11••,50]; W Smith, personal communication).
An alternate arachidonate-binding mode is observed in a
complex with the His207Ala mutant of COX-2 [11••]. The
carboxylate of arachidonate is hydrogen-bonded to Tyr385
and Ser530 and the ω-end projects toward the constriction
at Arg120, Tyr355, and Glu524 before bending up towardLeu531. This conformation is inconsistent with catalysisbut may correspond to an inhibitory conformation of substrate bound to enzyme.
The cyclooxygenase active sites of COX-1 and COX-2 are
very similar but there are subtle structural differences thatgive rise to functional differences between the two pro-
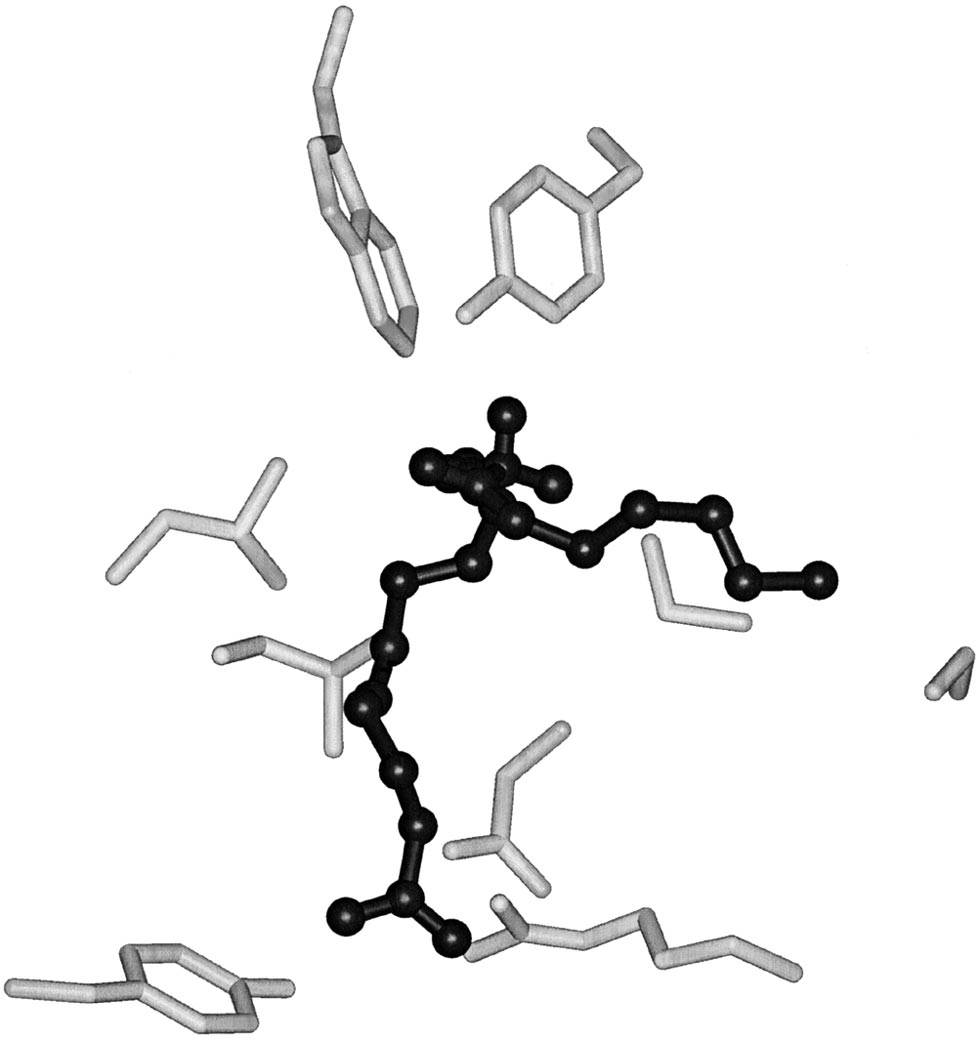
Model of arachidonic acid (black) bound to the active site of COX-1
teins. For example, aspirin acetylation of Ser530 of COX-1
(gray). Arg120, Tyr355 and Glu524 comprise the constriction thatseparates the bottom of the COX active site from the lobby in which
completely inhibits oxygenation of arachidonate by steri-
arachidonate first binds. The constriction must open to permit
cally blocking access to the top channel [51]. Aspirin
arachidonate access to the COX active site. Tyr385 sits adjacent to
acetylation of the corresponding residue in COX-2 abol-
the 13-pro(S) hydrogen of arachidonic acid and Trp387 facilitates
cyclization of the 11-peroxyl radical to form the cyclic peroxide.
Ser530 is the aspirin acetylation site and Gly533 is located near the
15-(R)-hydroxyeicosa-5,8,11,13-tetraenoic acid [52]. The
greater size of the cyclooxygenase active site in COX-2apparently allows insertion of arachidonate into the topchannel with an altered conformation of both the carboxyl
Conclusions
and ω-ends of the molecule; this leads to reversal in the
Recent work from several laboratories has provided
stereochemistry of oxygenation at C-15 [53•,54••].
important insights into the oxygenation of arachidonicacid by cyclooxygenases. These findings strongly support
A more fundamental difference between COX-1 and
the chemical mechanism of prostaglandin endoperoxide
COX-2 is in the binding of the carboxylate group of the
biosynthesis proposed over 30 years ago by Hamberg and
fatty acid substrate. The COX-1–arachidonate and
Samuelsson [2] and the biochemical mechanism of
COX-2–PGH2 crystal structures reveal ionic and hydrogen-
cyclooxygenase catalysis proposed 12 years ago by Ruf
bonding interactions with Arg120 and Tyr355, which are
and co-workers [8]. Reaching this level of understanding
located at a constriction point near the bottom of the
has been experimentally challenging because of the
cyclooxygenase active site and the top of the membrane-
short-lived nature of the substrate and enzyme-derived
binding domains of both proteins ([11••]; W Smith,
intermediates, the complex interaction between the
personal communication). As expected, mutations of
cyclooxygenase and peroxidase activities, and the
Arg120 of COX-1 significantly affect cyclooxygenase activ-
unusual kinetics of oxygenation that are complicated by
ity [55,56]. However, Arg120 mutations in COX-2 are much
less deleterious to its cyclooxygenase activity [57,58•]. Thissuggests that other interactions in the cyclooxygenase
We have begun to glimpse views of enzyme−substrate
active site are more important for binding arachidonate in
interactions that reveal the identity and role of residues
COX-2 than in COX-1. As a corollary, the carboxylate of
that control regiochemistry and stereochemistry of oxy-
arachidonate is not as important for its binding to COX-2 as
genation. Furthermore, we are beginning to appreciate
it is to COX-1. In support of this hypothesis, COX-2 oxy-
the subtle differences in structure between COX-1 and
genates the ethanolamide derivative of arachidonic acid
COX-2 that confer distinct substrate specificity and cat-
(anandamide) to the ethanolamide derivative of PGH
alytic function. The next few years should witness a
Mechanisms
more precise definition of enzyme–fatty-acid interac-
12. Tsai A-L, Palmer G, Xiao G, Swinney DC, Kulmacz RJ: Structural characterization of arachidonyl radicals formed by
tions for both enzymes. Because COX-2 oxygenates
prostaglandin H synthase-2 and prostaglandin H synthase-1
amide derivatives of arachidonic acid, it may be possible
reconstituted with mangano protoporphyrin IX. J Biol Chem 1998, 273:3888-3894.
to prepare fluorescent substrate analogs that enable real-
This paper follows earlier work from the same group demonstrating the
time monitoring of substrate binding and product release
ability of tyrosyl radicals generated by oxidation of wild-type or man-
[60]. This should provide a convenient approach for
ganese-substituted COX-2 to oxidize arachidonic acid to carbon-centeredradical derivatives. Tyrosyl radicals generated by oxidation of the
probing the involvement of individual residues in catal-
Tyr385Phe mutant of COX-2 do not react with arachidonic acid to form
ysis by both enzymes. Given the roles that COX
carbon-centered radicals, providing strong evidence for the identity of thetyrosyl radical generated from wild-type enzyme as derived of Tyr385.
enzymes play in lipid mediator biosynthesis, it is likely
13. Lassmann G, Odenwaller R, Curtis JF, Degray JA, Mason RP,
that these structural and functional differences will lead
Marnett LJ, Eling TE: Electron spin resonance investigation of
to important physiological consequences. tyrosyl radicals of prostaglandin H synthase. Relation to enzyme catalysis. J Biol Chem 1991, 266:20045-20055.
14. Kulmacz RJ, Palmer G, Wei C, Tsai AL: Reaction and free radical Acknowledgements kinetics of prostaglandin H synthase with manganese
I am grateful to J Prusakiewicz and GP Hochgesang for assistance with
protoporphyrin IX as the prosthetic group. Biochemistry 1994, 33:5428-5439.
some of the figures and to K Kozak for a critical reading. I am alsograteful to W Smith for helpful discussions related to the crystal
15. Degray JA, Lassmann G, Curtis JF, Kennedy TA, Marnett LJ, Eling TE,
structure of a COX-1–arachidonic acid complex. Work in the Marnett
Mason RP: Spectral analysis of the protein-derived tyrosyl radicals
laboratory has been supported by a research grant from the National
from prostaglandin H synthase. J Biol Chem 1992, 267:23583-23588.
16. Shi W, Hoganson CW, Espe M, Bender CJ, Babcock GT, Palmer G,
Kulmacz RJ, Tsai Al: Electron paramagnetic resonance and electron References and recommended reading nuclear double resonance spectroscopic identification and
Papers of particular interest, published within the annual period of review,
characterization of the tyrosyl radicals in prostaglandin H synthase 1. Biochemistry 2000, 39:4112-4121.
A detailed analysis of the tyrosyl radicals generated by oxidation of COX-1
that confirms earlier reports of the interconversion of radical signals by bond
rotation in the tyrosyl radical. ENDOR analysis suggests that the tyrosyl rad-ical signals detected in the presence of the COX inhibitor indomethacin are
Porter NA: Mechanisms for the autoxidation of polyunsaturated
derived from a different tyrosine than Tyr385. lipids. Acc Chem Res 1986, 19:262-268.
Shimokawa T, Kulmacz RJ, Dewitt DL, Smith WL: Tyrosine 385 of
Hamberg M, Samuelsson B: On the mechanism of the prostaglandin endoperoxide synthase is required for biosynthesis of prostaglandins E cyclooxygenase catalysis. J Biol Chem 1990, 265:20073-20076. 1 and F1α. J Biol Chem 1967, 242:5336-5343.
18. Gunther MR, Hsi LC, Curtis JF, Gierse JK, Marnett LJ, Eling TE,
Mason RP, Kalyanaraman B, Tainer BE, Eling TE: A carbon-centered
Mason RP: Nitric oxide trapping of the tyrosyl radical of free radical intermediate in the prostaglandin synthetase prostaglandin H synthase-2 leads to tyrosine iminoxyl radical and oxidation of arachidonic acid. Spin trapping and oxygen uptake nitrotyrosine formation. J Biol Chem 1997, 272:17086-17090. studies. J Biol Chem 1980, 255:5019-5022.
19. Goodwin DC, Gunther MH, Hsi LH, Crews BC, Eling TE, Mason RP,
Hecker M, Ullrich V, Fischer C, Meese CO: Identification of novel
Marnett LJ: Nitric oxide trapping of tyrosyl radicals generated arachidonic acid metabolites formed by prostaglandin H during prostaglandin endoperoxide synthase turnover: detection synthase. Eur J Biochem 1987, 169:113-123. of the radical derivative of tyrosine 385. J Biol Chem 1998, 273:8903-8909.
Xiao G, Tsai AL, Palmer G, Boyar WC, Marshall PJ, Kulmacz RJ:
Previous work has shown that nitric oxide quenches the tyrosyl radical deriv-
Analysis of hydroperoxide-induced tyrosyl radicals and
ative of COX-1 to form an iminoxyl radical. This article describes the isolation
lipoxygenase activity in aspirin-treated human prostaglandin H
of a single nitrated peptide from proteolytic digests of COX-1 following reac-
synthase-2. Biochemistry 1997, 36:1836-1845.
tion with arachidonic acid in the presence of nitric oxide sources and identi-fication of the nitrated residue as Tyr385. This indicates that Tyr385 is
Kwok P-Y, Muellner FW, Fried J: Enzymatic conversions of
oxidized to a tyrosyl radical trappable by nitric oxide. 10,10-difluoroarachidonic acid with PGH synthase and soybean lipoxygenase. J Am Chem Soc 1987, 109:3692-3698.
20. Dietz R, Nastainczyk W, Ruf HH: Higher oxidation states of prostaglandin H synthase. Rapid electronic spectroscopy
Dean AM, Dean FM: Carbocations in the synthesis of detected two spectral intermediates during the peroxidase prostaglandins by the cyclooxygenase of PGH synthase? reaction with prostaglandin G2. Eur J Biochem 1988, 171:321-328. A radical departure! Protein Sci 1999, 8:1087-1098.
21. Landino LM, Crews BC, Gierse JK, Hauser SD, Marnett LJ:
Karthein R, Dietz R, Nastainczyk W, Ruf HH: Higher oxidation states Mutational analysis of the role of the distal histidine and of prostaglandin H synthase. EPR study of a transient tyrosyl glutamine residues of prostaglandin-endoperoxide synthase-2 in radical in the enzyme during the peroxidase reaction. Eur J peroxidase catalysis, hydroperoxide reduction, and Biochem 1988, 171:313-320. cyclooxygenase activation. J Biol Chem 1997, 272:21565-21574.
Picot D, Loll PJ, Garavito RM: The X-ray crystal structure of the
22. Tsai AL, Wu G, Palmer G, Bambai B, Koehn JA, Marshall PJ,
membrane protein prostaglandin H 2 synthase-1. Nature 1994,
Kulmacz RJ: Rapid kinetics of tyrosyl radical formation and heme 367:243-249. redox state changes in prostaglandin H synthase-1 and -2. J Biol Chem 1999, 274:21695-21700.
10. Luong C, Miller A, Barnett J, Chow J, Ramesha C, Browner MF:
A rapid-freeze-quench and stopped-flow investigation of the formation of
Flexibility of the NSAID binding site in the structure of human
tyrosyl radicals and oxidized heme derivatives of COX-1 and COX-2. cyclooxygenase-2. Nat Struct Biol 1996, 3:927-933.
Formation of the first oxidized heme derivative precedes the detection of aprotein tyrosyl radical. This establishes the kinetic competence of the
11. Kiefer JR, Pawlitz JL, Moreland KT, Stegeman RA, Hood WF,
heme higher oxidation state in the generation of a tyrosyl radical that can
Gierse JK, Stevens AM, Goodwin DC, Rowlinson SW, Marnett LJ
oxidize arachidonic acid. Interestingly, the ferryl derivative produced coin-
et al.: Structural insights into the stereochemistry of the
cident with the tyrosyl radical is stable for COX-1 but rapidly decays to
cyclooxygenase reaction. Nature 2000, 405:97-111.
The first report of a structure of a COX enzyme with arachidonic acidbound. Arachidonic acid is bound in an inverted form that probably rep-
23. Smith WL, Eling TE, Kulmacz RJ, Marnett LJ, Tsai A: Tyrosyl radicals
resents an inhibitory complex with the enzyme. A second structure is
and their role in hydroperoxide-dependent activation and
reported that represents COX-2 with a molecule of the product PGH2
inactivation of prostaglandin endoperoxide synthase. Biochemistry
bound in the cyclooxygenase active site.
1992, 31:3-7. Cyclooxygenase mechanisms Marnett 551
24. Goodwin DC, Rowlinson SW, Marnett LJ: Substitution of tyrosine
and decay of heme higher oxidation states. Both enzymes react with
for the proximal histidine ligand to the heme of prostaglandin
hydroperoxides to form Intermediate I (Compound I) at comparable rates. endoperoxide synthase-2: implications for the mechanism of
However, the Intermediate I of COX-2 decays to Intermediate II (concomi-
cyclooxygenase activation and catalysis. Biochemistry 2000,
tant with tyrosyl radical formation) at much higher rates than the
39:5422-5432.
Intermediate I of COX-1. This accounts for the greater sensitivity of COX-2
Investigation of the possible role of ferrous COX-2 in cyclooxygenase activi-
to hydroperoxide-induced activation.
ty. Determination of the Fe3+–Fe2+ redox potentials of both COX-1 andCOX-2 suggests a significant thermodynamic barrier to reduction of Fe3+
42. Landino LM, Crews BC, Timmons MD, Morrow JD, Marnett LJ:
enzyme by a protein tyrosine residue. Site-directed mutation of the proximal
Peroxynitrite, the coupling product of nitric oxide and superoxide,
histidine of COX-2 to tyrosine produces an enzyme with high cyclooxyge-
activates prostaglandin biosynthesis. Proc Natl Acad Sci USA
nase activity after a prolonged lag phase. The activity of the His388Tyr
1996, 93:15069-15074.
mutant is also not consistent with reduction of Fe3+ enzyme because of the
43. Upmacis RK, Deeb RS, Hajjar DP: Regulation of prostaglandin H
difficulty of reducing heme with a tyrosinate ligand. The lag phase in
synthase activity by nitrogen oxides. Biochemistry 1999,
cyclooxygenase activity observed with the His388Tyr mutant is eliminated by
38:12505-12513.
addition of hydroperoxides in a manner that correlates to their ability to serveas peroxidase substrates.
44. Wadleigh DJ, Reddy ST, Kopp E, Ghosh S, Herschman HR:
Transcriptional activation of the cyclooxygenase-2 gene in
25. Kulmacz RJ, Wang LH: Comparison of hydroperoxide initiator endotoxin-treated RAW 264.7 macrophages. J Biol Chem 2000, requirements for the cyclooxygenase activities of prostaglandin H 275:6259-6266. synthase-1 and -2. J Biol Chem 1995, 270:24019-24023.
45. Chen CC, Chiu KT, Sun YT, Chen WC: Role of the cyclic AMP-
26. Chance B, Powers L, Ching Y, Poulos T, Schonbaum GR, Yamazaki I,
protein kinase A pathway in lipopolysaccharide-induced nitric
Paul KG: X-ray absorption studies of intermediates in peroxidase oxide synthase expression in RAW 264.7 macrophages. activity. Arch Biochem Biophys 1984, 235:596-611. Involvement of cyclooxygenase-2. J Biol Chem 1999, 274:31559-31564.
Defilippis MR, Murthy CP, Brotiman F, Weinraub D, Faraggi M, Klapper MH: Electrochemical properties of tyrosine phenoxy and
46. Marnett LJ, Wright TL, Crews BC, Tannenbaum SR, Morrow JD:
tryptophan indolyl radicals in peptides and amino acid analogues. Regulation of prostaglandin biosynthesis by nitric oxide is J Phys Chem 1991, 93:3416-3419. revealed by targeted deletion of inducible nitric-oxide synthase. J Biol Chem 2000, 275:13427-13430.
28. Koppenol WH: Oxyradical reactions: from bond-dissociation
Earlier work using in vitro experiments or in vivo pharmacological manipula-
energies to reduction potentials. FEBS Lett 1990, 264:165-167.
tions suggests a role for nitric oxide or a nitric-oxide-derived species in acti-
29. Tsai A-L, Kulmacz RJ, Wang J-S, Wang Y, Van Wart HE, Palmer G:
vating the cyclooxygenase activity of COX enzymes. This article reports that
Heme coordination of prostaglandin H synthase. J Biol Chem
macrophages isolated from mice with targeted deletions in the inducible form
1993, 268:8554-8563.
of nitric oxide synthase produce dramatically reduced levels of prostaglandinscompared with macrophages from wild-type animals. Similar results are
30. Page CC, Moser CC, Chen X, Dutton PL: Natural engineering
observed when comparing urinary prostaglandin levels from nitric oxide syn-
principles of electron tunnelling in biological oxidation-reduction.
thase-deficient mice to the levels from control mice. This provides direct evi-
Nature 1999, 402:47-52.
dence for a link between nitric oxide and prostaglandin biosynthesis, possiblymediated by cyclooxygenase activation by peroxynitrite.
31. Tang MS, Askonas LJ, Penning TM: Bromoacetamido analogs of indomethacin and mefenamic acid as affinity-labeling agents and
Marnett LJ, Rowlinson SW, Goodwin DC, Kalgutkar AS, Lanzo CA:
mechanistic probes for prostaglandin H2 synthase. Biochemistry Arachidonic acid metabolism by COX-1 and COX-2: mechanisms
1995, 34:808-815. of catalysis and inhibition. J Biol Chem 1999, 274:22903-22906.
32. Tang MS, Copeland RA, Penning TM: Detection of an Fe2+-
48. Rowlinson SW, Crews BC, Lanzo CA, Marnett LJ: The binding of protoporphyrin-IX intermediate during aspirin-treated arachidonic acid in the cyclooxygenase active site of mouse prostaglandin H2 synthase II catalysis of arachidonic acid to prostaglandin endoperoxide synthase-2 (COX-2): a putative 15-HETE. Biochemistry 1997, 36:7527-7534. L-shaped binding conformation utilizing the top channel region. J Biol Chem 1999, 274:23305-23310.
33. Cook HW, Lands WEM: Mechanism for suppression of cellular
Chemical principles of arachidonic acid oxygenation to PGG
biosynthesis of prostaglandins. Nature 1976, 260:630-632.
must bind in an extended conformation to generate a bicyclic peroxide with
34. Strieder S, Schaible K, Scherer H-J, Dietz R, Ruf HH: Prostaglandin trans-dialkyl substitution. Modeling of this conformation into the COX-2
endoperoxide synthase substituted with manganese
active site suggests the ω-end of the fatty acid is bound in a top channel
protoporphyrin IX. Formation of a higher oxidation state and its
close to Gly533. Mutation of Gly533 to Ala, Val, or Leu drastically reduces
relation to cyclooxygenase reaction. J Biol Chem 1992,
cyclooxygenase activity with arachidonic acid as substrate but not with
267:13870-13878.
unsaturated fatty acids with fewer carbons at their ω-ends. These results areconsistent with the proposed model for arachidonic acid binding. This was
35. Odenwaller R, Maddipati KR, Marnett LJ: Detection of a higher
subsequently confirmed by crystallographic analysis of COX-1 and COX-2
oxidation state of manganese-prostaglandin endoperoxide
complexes with arachidonic acid and PGH2 (see text). synthase. J Biol Chem 1992, 267:13863-13869.
49. Hsi LC, Tsai A, Kulmacz RJ, English DG, Siefker AO, Otto JC,
36. Kulmacz RJ, Lands WEM: Requirements for hydroperoxides by the
Smith WL: Trp387 and the putative leucine zippers of PGH cyclooxygenase and peroxidase activities of prostaglandin H synthases-1 and -2. J Lipid Mediat 1993, 6:131-138. synthase. Prostaglandins 1983, 25:531-540.
50. Thuresson ED, Lakkides KM, Smith WL: Different catalytically
Nugteren DH, Hazelhof E: Isolation and properties of intermediates competent arrangements of arachidonic acid within the in prostaglandins biosynthesis. Biochim Biophys Acta 1973, cyclooxygenase active site of prostaglandin endoperoxide H 326:448-461. synthase-1 lead to the formation of different oxygenated products. J Biol Chem 2000, 275:8501-8507.
38. Hamberg M, Svensson J, Wakabayashi T, Samuelsson B: Isolation and structure of two prostaglandin endoperoxides that cause
51. Shimokawa T, Smith WL: Prostaglandin endoperoxide synthase. platelet aggregation. Proc Natl Acad Sci USA 1974, 71:345-349. The aspirin acetylation region. J Biol Chem 1992, 267:12387-12392.
39. Tsai A-L, Wu G, Kulmacz RJ: Stoichiometry of the interaction of prostaglandin H synthase with substrates. Biochemistry 1997,
52. Lecomte M, Laneuville O, Ji C, Dewitt DL, Smith WL: Acetylation of 36:13085-13094. human prostaglandin endoperoxide synthase-2 (cyclooxygenase-2) by aspirin. J Biol Chem 1994, 269:13207-13215.
40. Markey CM, Alward A, Weller PE, Marnett LJ: Quantitative studies of hydroperoxide reduction by prostaglandin H synthase. J Biol
53. Rowlinson SW, Crews BC, Goodwin DC, Schneider C, Gierse JK,
Chem 1987, 262:6266-6279.
Marnett LJ: Spatial requirements for 15-HETE synthesis within the cyclooxygenase active site of murine COX-2: why acetylated
41. Lu G, Tsai AL, Van Wart HE, Kulmacz RJ: Comparison of the COX-1 does not synthesize 15-(R)-HETE. J Biol Chem 2000, peroxidase reaction kinetics of prostaglandin H synthase-1 and -2. 274:6586-6591. J Biol Chem 1999, 274:16162-16167.
Aspirin-acetylated COX-2 makes 15-(R)-HETE, whereas aspirin-acetylated
Previous work from several groups established that cyclooxygenase activi-
COX-1 does not. Site-directed mutants that convert COX-2 residues to
ty of COX-2 is activated at significantly lower hydroperoxide concentrations
COX-1 residues near the bottom and the top of the cyclooxygenase active
than COX-1. This article reports a detailed kinetic analysis of the formation
site abolish the ability of acetylated COX-2 to make 15-(R)-HETE. Mechanisms
54. Schneider C, Brash AR: Stereospecificity of hydrogen abstraction
58. Rieke CJ, Mulichak AM, Garavito RM, Smith WL: The role of arginine in the conversion of arachidonic acid to 15R-HETE by aspirin- 120 of human prostaglandin endoperoxide H synthase-2 in the treated cyclooxygenase-2. Implications for the alignment of interaction with fatty acid substrates and inhibitors. J Biol Chem substrate in the active site. J Biol Chem 2000, 275:4743-4746.
1999, 274:17109-17114.
Arachidonic acid is converted into PGG2 following removal of the 13-pro(S)
Arg120 is the only positively charged residue in the cyclooxygenase active
hydrogen. The stereochemistry of oxygenation at C-15 is (S). The isolation of
site and ion-pairs to the carboxylates of arachidonic acid and arylacetic acid
15-(R)-HETE from aspirin-acetylated COX-2 suggests that the stereochem-
inhibitors. Mutation of Arg120 of COX-1 to Gln dramatically increases the
istry of hydrogen removal from arachidonic acid may be reversed from that
Km of the mutant when acting on arachidonic acid. In this follow-up article,
observed with wild-type enzyme. By synthesizing enantiospecifically labeled
the authors demonstrate that a similar mutant of COX-2 has little or no effect
arachidonic acid and reacting it with acetylated COX-2, the authors were
on the Km for arachidonate. This suggests that ionic interactions are less
able to demonstrate that the 13-pro(S) hydrogen is removed. This suggests
important for binding substrate in COX-2 than in COX-1.
that the (R)-stereochemistry of oxygenation at C-15 observed with acetylatedCOX-2 results from O
59. Yu M, Ives D, Ramesha CS: Synthesis of prostaglandin E
2 trapping of an altered conformation of a putative
pentadienyl radical leading to 15-HETE synthesis. ethanolamide from anandamide by cyclooxygenase-2. J Biol Chem 1997, 272:21181-21186.
55. Mancini JA, Riendeau D, Falgueyret J-P, Vickers PJ, O’Neill GP:
Arginine 120 of prostaglandin G/H synthase-1 is required for the
60. Lanzo CA, Sutin J, Rowlinson SW, Talley J, Marnett LJ: Fluorescence inhibition by nonsteroidal anti-inflammatory drugs containing a quenching analysis of the association and dissociation of a carboxylic acid moiety. J Biol Chem 1995, 270:29372-29377. diaryheterocycle to cyclooxygenase-1 and cyclooxygenase-2: the dynamic basis of cyclooxygenase-2 selectivity. Biochemistry 2000,
56. Bhattacharyya DK, Lecomte M, Rieke CJ, Garavito RM, Smith WL:
39:6228-6234. Involvement of arginine 120, glutamate 524, and tyrosine 355 in the binding of arachidonate and 2-phenylpropionic acid inhibitors to the cyclooxygenase active site of ovine prostaglandin endoperoxide H synthase-1. J Biol Chem 1996, 271:2179-2184. Now in press The work referred to in the text as ‘W Smith, personal communication’ is now
Greig GM, Francis DA, Falgueyret JP, Ouellet M, Percival MD, Roy P,
Bayly C, Mancini JA, O’Neill GP: The interaction of arginine 106 of human prostaglandin G/H synthase-2 with inhibitors is not a
61. Malkowski MG, Ginell SL, Smith WL, Garavito RM: Structure of universal component of inhibition mediated by nonsteroidal anti- prostaglandin synthase complexed with arachidonic acid. Science inflammatory drugs. Mol Pharmacol 1997, 52:829-838.
types aggregate in QTL regions, combined withas an iv bolus to determine the contribution of nitric14. E. S. Lander et al. , Genomics 1 , 174 (1987). physiological profiling, provides a novel ap-oxide to basal renal vascular tone. After 10 min of15. J. Loscalzo, G. Welch, Progr. Cardiovasc. Dis. 38 , 87 equilibration, a repeat infusion of the same two doses ofproach to facilitate the
The Research Literature: Academic MonitoringBrophy and Good (1986) made the following observation on the importance and complexity Elitist critics often undervalue teaching, or even suggest that anyone can teach. ("Those who can, do; those who can’t, teach.") The data reviewed here refute this myth well. Although it may be true that most adults could survive in the classroom, it is not
 Cyclooxygenase mechanisms Marnett 549
Cyclooxygenase mechanisms Marnett 549